ALTE DOCUMENTE |
PRINCIPALELE PROCESE SI RELATII IN VIATA ARBORETULUI
Spre deosebire de arbore a carui existenta este finita, adica are un inceput si un sfarsit, arboretul, prin procesul de regenerare naturala, se poate mentine o foarte lunga perioada de timp (pana la inlocuirea lui succesionala prin alt tip de arboret). Dar in aceasta lunga existenta arboretul este intr-o continua schimbare printr-o serie de procese care se desfasoara continuu sau periodic . Aceste procese sunt:
procesul de regenerare naturala a arboretului,
procesul de formare a starii de masiv,
procesul de eliminare naturala a arborilor,
procesul de diferentiere a arborilor pe inaltime,
procesul de acumulare de lemn in arboret,
procesul de elagare naturala si de modificare a formei trunchiului
arborilor.
5.1. PROCESUL DE REGENERARE NATURALA A ARBORETULUI
Regenerarea naturala a arboretului este procesul de formare a unei generatii noi de arbori fie din samanta cazuta la sol din arborii generatiei vechi, fie din lastarii (drajonii) proveniti din acesti arbori, fie atat din samanta cat si din lastarii acestora. Este vorba despre un proces natural care insa in paduri cultivate poate fi dirijat de silvicultor.
Desigur prin procesul de regenerare se produc si profunde modificari in intreaga biocenoza forestiera dar silvicultorul intervine direct doar asupra arboretului si numai in unele cazuri si asupra altor componente biocenotice (arbusti, ierburi) sau a solului. In urma regenerarii arboretului se reconstituie pe cale naturala intreaga biocenoza forestiera, fara interventia silvicultorului.
Procesul de regenerare naturala poate fi continuu daca arboretul este plurien, format din arbori de toate varstele din care, permanent, cei de varste inaintate dispar iar cei de varste mijlocii fructifica si insamanteaza terenul devenind astfel liber. Dar procesul de regenerare naturala poate fi si periodic, in cazul cand arboretul este echien si cand disparitia arborilor batrani si nasterea noului arboret se produce intr-o perioada relativ scurta de timp (pana la 20-30 de ani) iar urmatoarea schimbare de generatii intervine dupa un secol sau mai mult.
Regenerarea naturala se poate produce din samanta arborilor din vechiul arboret, sub masivul acestuia dar si in teren deschis, in care samanta ajunge din arboretele vecine.
Dar regenerarea naturala se poate produce si vegetativ, din lastarii porniti din tulpina sau radacina arborilor (drajoni), in cazul vatamarii acestora prin factori biotici (consum) sau abiotici (rupere de vant, zapada). In conditii naturale, regenerarea vegetativa este mai putin frecventa decat cea din samanta. Capacitate de regenerare vegetativa nu au toti arborii ci, in majoritate, cei de foioase la care se pot forma muguri dorminzi sau adventivi. Cele mai multe specii de arbori rasinosi nu au aceasta insusire.
5.1.1. Regenerarea naturala din samanta
In procesul de regenerare naturala din samanta prima etapa importanta este fructificatia arborilor. Aceasta se produce, in arboret, la varste destul de mari, la arbori la care se incetineste cresterea in inaltime iar, prin eliminarea naturala, coroanele celor ramasi capata mai multa lumina.
Fructificatia in arboret nu se produce anual decat la putine specii, ci periodic la interval de cativa ani. Cauza este dezvoltarea limitata a coroanelor arborilor in masiv si ca urmare producerea de mai putina biomasa din care se pot forma flori, fructe, seminte. In masiv exista deci o periodicitate a fructificatiei, diferita pe specii.
Varsta la care incepe fructificatia arborilor in masiv si periodicitatea acesteia este data in tabelul 5.1.
Tabelul 5.1.
Varsta inceperii si periodicitatea fructificatiei la cateva specii de arbori - Dupa I. Florescu 1981
Specia |
Inceperea fructificatiei la arbori izolati…ani |
Inceperea fructificatiei la arborii in masiv…ani |
Periodicitatea fructificatiei arborilor in masiv…ani |
|
Molid | |||
|
Brad | |||
|
Fag | |||
|
Gorun | |||
|
Stejar pedunculat | |||
|
Carpen | |||
|
Tei, paltini, frasini | |||
|
Larice, pin silvestru | |||
|
Salcam |
Intensitatea (abundenta) fructificatiei poate sa fie foarte diferita atat la diferite specii cat si in ani diferiti la aceeasi specie. Fructifica de regula abundent speciile de amestec (carpenul, teii, frasinii, ulmii).
In unii ani arborii pot fructifica abundent (ani de samanta) in altii putin abundent (ani de stropeli in fructificatie), in altii deloc.
Pentru silvicultor sunt de luat in seama si anii de stropeli care pot furniza samanta, in completare, pentru regenerarea naturala dar si pentru semanarea in pepiniere.
Asa cum rezulta din tabelul 5.1. varsta inceperii fructificatiei in masiv la speciile principale este destul de mare (60-70 de ani la brad, fag, gorun). Mai devreme incepe fructificatia la speciile de amestec si la rasinoasele heliofite si mai ales la salcam. Periodicitatea este de asemenea mai mare la speciile principale, mai mica la speciile de amestec. Aceste diferente sunt legate si de longevitatea speciilor.
Este de relevat si faptul ca arborii regenerati vegetativ fructifica mai devreme decat cei regenerati din samanta.
In arboret, arborii nu fructifica la fel. Arborii dominanti, dar in special cei predominanti, produc majoritatea semintelor deoarece au coroana bine dezvoltata si luminata, cei 353e42d codominanti fructifica putin , cei dominati deloc.
Productia de samanta este diferita pe specii ( tabelul 5.2.).
Tabelul 5.2.
Productia de samanta in unele arborete (dupa I. Florescu 1996 din Negulescu si Ciumac 1959).
|
Arboretul |
Productia medie de samanta kg/an/ha |
|
Molidis | |
|
Faget | |
|
Stejaret | |
|
Laricet | |
|
Pinet |
Cunoasterea de catre silvicultor atat a varstei de incepere a fructificatiei, cat si a periodicitatii si intensitatii (abundentei) ei este foarte importanta pentru fixarea ciclului de productie, alegerea momentului pentru taierile de regenerare, producerea de puieti in pepiniera etc.
De aceea este foarte importanta evaluarea anuala a fructificatiei. Aceasta se poate face la inflorire prin apreciere (bogata, medie, slaba) sau prin numararea florilor pe lujeri de proba. Dar se face si pe parcursul formarii semintelor sau la diseminare atat prin apreciere, dar mai precis prin culegerea fructelor sau semintelor din arbori de proba, sau recoltarea acestora de sub asemenea arbori si apoi prin numarare sau cantarire.
Calitatea semintelor nu este aceeasi la diferite specii si in diferiti ani. Calitatea semintelor se evalueaza dupa capacitatea lor de germinare si de formare a puietilor viabili. Calitatea depinde de specie, de bonitatea stationala, de cantitatea de rezerve nutritive pe care o contine samanta, de starea ei de sanatate. In anii cu clima nefavorabila, cu atacuri de insecte sau de ciuperci, numai o cantitate redusa de samanta este viabila, desi fructificatia poate fi abundenta.
Repausul vegetativ al semintelor. La multe specii de arbori semintele au un repaus vegetativ inainte de a fi capabile de germinare. Repausul poate fi de cateva zile la salcii, plopi, ulmi, dar de cateva luni sau chiar mai mult de un an (frasini, tei, carpen). De aceea chiar in anii in care nu exista fructificare, pot aparea puieti pentru ca exista o rezerva de seminte in sol. Chiar in cazul semintelor la care repausul vegetativ este scurt ramane o cantitate de samanta care germineaza mai tarziu (cazul fagului la care semintisul poate apare si dupa un an sau doi de la inlaturarea arboretului batran). Sunt specii la care intarzierea germinarii este cauzata de tegumentul gros al semintelor (salcam, gladita, zambru).
Germinarea semintelor si formarea plantulelor (puietilor) se produce daca sub masiv sau in teren descoperit se realizeaza conditii de umiditate si caldura, la unele specii si de lumina, favorabile. Germinarea se poate produce chiar dupa diseminare in toamna (fag, stejar) dar mai frecvent primavara.
Puterea de rasarire se exprima prin procentul de plantule rasarit din totalul semintelor cazute. Acest procent nu este acelasi cu procentul de germinare care poate fi mai mare pentru ca unele seminte germineaza dar nu ajung sa formeze puieti. De exemplu la molid daca samanta a cazut pe stratul gros de humus brut si a germinat iar radacinuta pornita din samanta nu ajuns la solul mineral plantula nu se va mai forma.
Formarea semintisului se poate produce simultan, pe toata suprafata arboretului batran, intr-un an de fructificatie abundenta, sau in etape, pe suprafete partiale, in mai multi ani cu fructificatii mai reduse.
In formarea si dezvoltarea puietilor care vor constitui semintisul, importante sunt conditiile de lumina, de caldura, de apa si substante din sol dar si de litiera necromasa nedescompusa care acopera solul. La cele mai multe specii de arbori conditiile cele mai favorabile pentru formarea si dezvoltarea puietilor este in mediul microclimatic al padurii, cu lumina suficienta (dupa cerintele fiecarei specii) dar in atmosfera umeda, calda, fara pericol de ingheturi, pe soluri neintelenite, neacoperite de un strat gros de litiera nedescompusa, suficient de afanate, cu rezerve suficiente de apa. La molid de exemplu, in cazul solurilor cu humus brut, puietii se instaleaza doar pe taluzuri, pe solul mineral, sau pe lemnul putred din trunchiurile cazute pe sol.
Formarea semintisului se incheie odata cu dezvoltatea suficienta a puietilor pentru ca micile lor coroane sa vina in contact si sa se formeze starea de masiv. Odata format masivul de puieti, se poate considera incheiat procesul de regenerare a arboretului si incepe alt proces – de eliminare naturala.
Dinamica numarului de puieti, pana la formarea starii de masiv, depinde mai mult de factorii nefavorabili biotici sau abiotici deoarece in aceasta faza competitia intre puieti nu este inca declansata decat in palcurile prea dese.
Dar chiar in lipsa competitiei, numarul de puieti scade din primavara pana in toamna chiar in primul an de viata (tabelul 5.3.).
Tabel 5.3.
Variatia numarului de puieti in primul an de viata intr-un faget natural dupa V. Stanescu si colaboratorii 1970.
|
Specificare |
V |
V |
VI |
VII |
VIII |
IX |
X |
XI |
|
Numar de puieti la ha (mii bucati) |
Rezistenta puietilor si a semintisului sub masiv depinde de cerintele de lumina dar si de apa al puietilor, foarte diferite pe specii, dar diferite si in functie de varsta arborilor. La puieti, de exemplu, cerintele fata de lumina pot fi mai reduse decat la arbori maturi.
Cerinte mari fata de lumina au puietii de stejar pedunculat, mesteacan, pin silvestru, larice, cerinte ceva mai reduse au puietii de molid, gorun, garnita, tei, frasini, paltini etc, cerinte foarte reduse au puietii de fag, brad, carpen, tisa. Puietii cu cerinte mari de lumina rezista sub masiv doar 1-2 ani, cei cu cerinte ceva mai reduse 2-4 ani, cei cu cerinte reduse 10-15 ani si chiar pana la 50 de ani. In cazul cand fluxul de lumina se amplifica puietii isi pot reactiva cresterea daca acest interval de timp nu este depasit. Altfel, la puieti nu se mai produce cresterea anuala sau aceasta este foarte redusa iar, dupa un timp, puietii se usuca, incetand sinteza glucidelor necesare consumului curent.
Puietii care au cerinte mari fata de lumina se pot forma si dezvolta si in teren deschis dar cei cu cerinte reduse au nevoie, un timp destul de indelungat, de protectia arboretului batran atat impotriva insolatiei prea puternice cat si impotriva ingheturilor. Acestea nu se produc sub adapostul creat de arborii batrani pentru ca, coroanele lor impiedica radiatia calorica nocturna prea puternica a solului, din cauza careia se produce inghetul la sol.
Cunoasterea cerintelor fata de lumina a puietilor este necesara pentru silvicultor pentru dirijarea, prin taieri, a conditiilor de lumina din arboretul in curs de regenerare, in vederea asigurarii, mentinerii si dezvoltarii suficient de active a puietilor sub masiv, pana la punerea lor in lumina plina prin eliminarea completa a arboretului batran.
Regenerarea naturala din samanta are multe avantaje in padurea cultivata. In primul rand este un proces natural in care se conserva, integral, biodiversitatea arborilor in generatia noua. In al doilea rand prin regenerarea din samanta se dezvolta arbori longevivi, cu capacitate productiva normala pentru specie, pana la varste mari, cu trunchiuri de forme mai bune si lemn de dimensiuni mai mari si de calitate mai buna. Arbori sunt mai rezistenti la boli (in special micotice) si la daunatori, ca si la factori abiotici.
Din punct de vedere economic regenerarea naturala este mai ieftina decat cea artificiala dar necesita, de regula, un timp mai indelungat pana se obtine un semintis suficient pe intreaga suprafata.
In silvicultura ecologica este principala modalitate de regenerare a arboretelor acceptata si promovata astazi din ce in ce mai mult.
Regenerarea naturala din samanta a arboretelor pluriene. Fata de modul de regenerare a arboretelor echiene sau relativ echiene, regenerarea in padurile pluriene, in care coexista arbori de toate varstele, prezinta deosebiri importante.
Deoarece in arboretele pluriene arborii batrani sunt de regula putini la numar si imprastiati pe suprafata, disparitia lor creeaza ochiuri cu lumina ceva mai multe dar pe suprafete mici, dispersate. In aceste locuri se poate instala un semintis si dezvolta in continuare un palc de arbori tineri. Disparitia arborilor batrani producandu-se continuu si procesul de regenerare are caracter continuu, spre deosebire de cel din padurile echiene care are caracter periodic. Astfel, in asemenea arborete, semintisul se poate gasi peste tot, pe mici suprafete, alaturi de palcuri de arbori tineri, de arbori maturi, de arbori batrani, ca si de arbori batrani in curs de uscare.
Desigur in arborete pluriene se pot regenera numai speciile a caror puieti pot rezista la umbra un timp mai indelungat pentru ca ochiurile create prin disparitia arborilor batrani sunt de dimensiuni reduse si mai mult sau mai putin umbrite lateral(este vorba de brad, fag, intr-o anumita masura si molid).
5.1.2. Regenerarea naturala vegetativa
Se poate produce numai la speciile de arbori care au muguri dorminzi sau pot forma muguri adventivi si se dezvolta in cazul ruperii naturale sau a taierii trunchiului. Lastarii formati din muguri dorminzi (proventivi) se numesc lastari proventivi, cei formati din muguri adventivi sunt lastari adventivi. Lastarii proventivi sunt calitativ mai buni si mai rezistenti la vatamari fiind stadiali mai tineri. Cei adventivi sunt calitativ mai slabi, se desprind usor de trunchi, sunt frecvent vatamati de putregai.
Regenerarea vegetativa se produce prin lastari proveniti fie din cioata trunchiului fie din radacina (caz in care lastarii respectivi se numesc drajoni). Dar regenerarea vegetativa se poate produce si din lastari detasati din arbori (de exemplu la salcii, plopi). Acest mod de regenerare se practica de silvicultori prin confectionarea si plantarea de butasi.
Este de mentionat ca la unele specii de arbori regenerarea vegetativa din drajoni se produce chiar daca arborele nu este vatamat, constituind un mod de ocupare a terenului (asa se intampla la plopul alb si tremurator, la ulm, la salcam). La unele specii regenerarea vegetativa se produce si prin lastari subterani, care se dezvolta pe o anumita lungime sub pamant, iar apoi, la o distanta de arborele batran, apar si se dezvolta deasupra solului (asa se intampla la stejarul pufos, posibil la garnita si tei argintiu) in conditii stationale nefavorabile.
Lastarii subterani, drajonii cat si lastari ce se formeaza la baza trunchiului se hranesc la inceput din radacina arborelui batran dar isi pot forma apoi radacini proprii. Daca lastarii se formeaza mai sus, ei se hranesc numai pe seama radacinii arborelui disparut.
Fiind hraniti la inceput de o radacina mare, toti lastarii si drajonii cresc mai repede decat puietii formati din samanta. Dar dupa cativa ani vigoarea de crestere scade iar productivitatea se reduce.
Lastaresc bine, chiar la varste mai mari (dar nu peste 30-40 de ani), stejarul, gorunul, teii, carpenul, jugastrul, aninul, plopii, salciile. Lastaresc bine dar numai la varste tinere fagul, mesteacanul, salcamul.
Drajoneaza bine plopul alb, plopul tremurator, cenusarul, salcamul, ulmii. Drajonii pot apare la distante relativ mari de cioata sau arbore (la salcam la 10-15 m, la cenusar pana la 30-35 m).
Dezvoltarea lastarilor si drajonilor fiind foarte activa din primul an, starea de masiv a lastarisurilor se realizeaza rapid. Daca in suprafata exista si puieti din samanta, care in primii ani se dezvolta mult mai incet decat lastarii sau drajonii, puietii din samanta vor fi eliminati.
Lastarirea este mai viguroasa in statiuni de bonitate ridicata. Daca primii lastari sunt distrusi (de exemplu prin scoaterea arborilor doborati sau a resturilor de exploatare), se poate forma alta serie de lastari, dar de vitalitate redusa.
Regenerarea vegetativa se poate produce la orice varsta a arborilor. De exemplu la puieti, din cauza umbrei, a secetei, a vatamarii prin alti factori, se poate usca tulpina (fenomen de autorecepare). Dar de la baza tulpinei poate porni un lastar care sa refaca puietul.
La arborii care isi pierd o parte din coroana sau chiar intreaga coroana prin uscare, fenomen numit „coronare”, din mugurii existenti pe tulpina se pot forma asa numitele „craci lacome”. Acestea tind sa refaca, macar partial coroana pentru ca volumul de substante organice produs prin fotosinteza sa nu scada. Fenomenul este frecvent la stejari, uneori chiar daca coroana nu se usuca dar trunchiul este prea luminat. In consecinta lemnul capata noduri si calitatea acestuia scade.
5.2. PROCESUL DE FORMARE A STARII DE MASIV
Atunci cand coroanele puietilor, proveniti din samanta (natural sau artificial) sau a lastarilor proveniti din regenerarea vegetativa, ajung sa se atinga formand un invelis neintrerupt sau aproape neintrerupt, se realizeaza cea mai importanta caracteritica a noului arboret – starea de masiv. Aceasta stare, care defineste de fapt padurea, se va mentine apoi pana la imbatranirea arboretului si rarirea lui naturala, prin disparitia arborilor batrani. Starea de masiv isi va pune pecetea pe dezvoltarea ulterioara a arborilor si pe structura viitorului arboret, dar si a intregii biocenoze ca si asupra statiunii.
In cazul arboretului echien, disparitia in masa a arborilor batrani determina intreruperea temporara a starii de masiv pe suprafete mari si refacerea ei atunci cand semintisul acopera din nou toata suprafata. In cazul arboretului plurien, starea de masiv este permanenta deoarece procesul de formare a semintisului este continuu si se produce pe mici suprafete, acolo unde dispar arborii foarte batrani.
In conditii favorabile pentru biocenozele forestiere, adica in climate suficient de calde si de umede, existenta normala a arborilor este sub forma de masiv. Este adevarat ca in asemenea climate arborii pot creste si izolat dar aceasta este o exceptie. Pentru ca, asa cum rezulta din teoria genezei padurii, din cauza producerii unei mari cantitati de samanta, spatiul liber este repede ocupat, in intregime de semintisuri abundente, sub forma de masiv, care se va mentine apoi de-a lungul vietii arboretului. Doar in conditii putin favorabile cresterii arborilor, in climate prea reci, prea uscate unde arborii au longevitate redusa si fructificatie putin abundenta, rara, arborii cresc izolat sau sub forma de raristi deci fara a forma un masiv.
Formarea starii de masiv se produce mai repede (in 2-4 ani) la speciile cu crestere rapida la inceput (salcam, larice, plopi salcii) dar destul de incet (in 7-15 ani) la speciile cu crestere inceata in primii ani (stejari, fag, brad). In lastarisuri provenite din regenerare vegetativa starea de masiv se instaleaza repede (2-3 ani).
Rapiditatea formarii starii de masiv depinde si de bonitatea stationala: inchegarea arboretului se face mai curand in statiuni de bonitate superioara decat in cele de bonitate inferioara.
Consecintele formarii starii de masiv asupra dezvoltarii ulterioare a arborilor si a arboretului sunt deosebite.
Prima consecinta este declansarea competitiei intre puieti pentru spatiu si factorii vitali (lumina, apa, elementele nutritive). Ca rezultat incepe, foarte activ, procesul de eliminare naturala a arborilor care se va desfasura de-a lungul intregei existente a arboretului. (figura 5.1.).
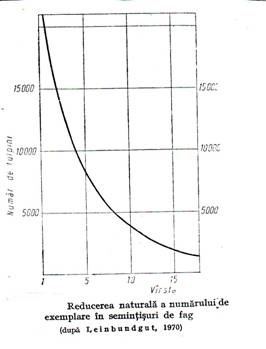
Figura 5.1.
A doua consecinta este diferentierea arborilor de aceeasi varsta dupa inaltime, grosime, marimea si forma coroanei. Se naste astfel o structura caracteristica a arboretului.
A treia consecinta este elagarea naturala a trunchiului arborilor, adica uscarea si apoi desprinderea de trunchi a ramurilor inferioare lipsite de lumina. Acest proces deosebeste transant arborii crescuti in masiv de cei crescuti izolat si are o deosebita importanta din punct de vedere a calitatii lemnului si a valorii acestuia.
Dar, consecintele formarii starii de masiv a arboretului sunt hotaratoare si pentru formarea intregii biocenoze, prin selectarea doar a speciilor de plante, animale, microorganisme care pot trai in mediul special creat in masivul si sub masivul arborilor, dar si pentru formarea biotopului, prin modificarea radicala a fluxurilor de radiatie si apa si a circuitelor de elemente chimice in statiunile ocupate de padure.
5.3. PROCESUL DE ELIMINARE NATURALA A ARBORILOR IN MASIV
Odata cu formarea starii de masiv, declansarea competitiei intre puietii in crestere, care au nevoie de mai mult spatiu si resurse abiotice, determina inceperea indelungatului proces de eliminare naturala a arborilor. Acest proces, foarte intens la inceput, se incetineste cu varsta. Daca initial in semintis pot exista milioane de puieti, in arboretele batrane raman doar 200-400 arbori. Desfasurarea procesului este ilustrata prin curba scaderii numarului de arbori cu varsta.
Tabelul 5.4.
Reducerea numarului de puieti de fag cu varsta in statiune de bonitate superioara - dupa Assmann, modificat
|
Varsta Ani |
Multimea de arbori |
Diametrul mediu cm |
|
|
numar/ha | |||
|
| |||
Este de relevat ca cea mai intensa eliminare este in primii 10 ani cand 88% din puieti dispar. Dupa 40 de ani se produce injumatatirea numarului de puieti la fiecare 20 de ani.
Intensitatea atat de mare a acestui proces este cauzata de talia mare a arborilor care face ca spatiul vital minim necesar sa creasca puternic cu varsta, pe masura ce se dezvolta coroana si radacina.
Reducerea numarului de arbori cu varsta la speciile autohtone si la cateva specii alohtone este redata in tabelele de productie romanesti (Giurgiu si colab. 1972) (figura 5.2.).
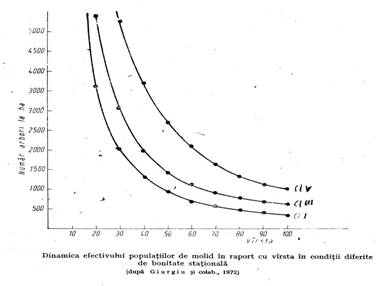
Figura 5.2.
Procesul de eliminare naturala depinde de factori interni ai arborilor (zestrea genetica) si de factori externi (relatiile biocenotice, complexul de factori stationali, micromozaicul stational). In legatura cu acest proces se vorbeste de un aparent paradox: cu cat statiunea este de bonitate mai ridicata, intensitatea eliminarii este mai mare. S-ar parea ca in asemenea conditii favorabile ar putea coexista un numar mai mare de arbori, dar nu este asa deoarece, tocmai datorita acestor conditii, arborii au coroane si radacini mai mari si avand nevoie de mai mult spatiu, eliminarea este mai puternica. Si acest fenomen este ilustrat de tabelele de productie in care, la aceeasi varsta, numarul de arbori din arborete de clasa I de productie este de 2-3 ori mai mic decat in cele de clasa V.
Procesul de eliminare naturala este, in fond, si un proces aspru de selectie naturala care elimina exemplarele mai slab dezvoltate, vatamate, genetic mai slab adaptate la mediul abiotic sau la competitie. Prin aceasta selectie populatia, devine din ce in ce mai bine adaptata la mediul abiotic sau la competitie. Este adevarat ca prin selectia naturala nu se selecteaza in toate cazurile arborii cei mai buni din punct de vedere silvicultural ci arborii cei mai vigurosi si rezistenti la impacturi. De aceea, interventia silvicultorului in procesul de eliminare naturala devine necesara pentru a favoriza supravietuirea arborilor cu trunchiuri valoroase din punct de vedere economic si crearea de structuri ale arboretului cat mai favorabile dezvoltarii acestora.
In arboretele pure, eliminarea se produce doar prin competitie intrapopulationala, care este orientata spre mentinerea si perfectionarea speciei. In arboretele amestecate intervine insa si competitia intre populatii, care poate fi mai intensa, iar uneori chiar eliminatorie, pentru una din populatii. Astfel in amestecurile de specii de lumina si de umbra (gorun + fag) sau de specii repede si incet crescatoare (carpen + stejar pedunculat) este posibila eliminarea unei populatii. Totusi, asa cum arata situatiile reale, chiar in asemenea cazuri, exista mecanisme care fac posibila coexistenta populatiilor concurente.
Eliminarea naturala este mai intensa si incepe mai repede la speciile de lumina si la cele repede crescatoare decat la speciile de umbra si cele incet crescatoare.
Eliminarea naturala se produce si in arboretele pluriene. Dar, spre deosebire de cele echiene in care numarul arborilor scade puternic cu varsta, in arboretele pluriene numarul de arbori ramane ± constant. Acest lucru este posibil pentru ca eliminarea se produce simultan la toate varstele si cu intensitati diferite in functie de varsta. Raman la fiecare varsta numai atatia arbori cat permite spatiul vital restrans ce revine varstei respective, astfel ca pe ansamblu numarul de arbori nu se schimba.
Silvicultorul trebuie sa fie bine familiarizat cu felul cum se produce eliminarea naturala in fiecare populatie ecologica, atat in arborete pure cat si amestecate si in statiuni de bonitate diferita. Intensitatea interventiilor de conducere a arboretelor, adica eliminare artificiala, trebuie sa fie destul de apropiata de cea naturala la care populatia este adaptata. Intensitati mai mici sau mai mari se pot adopta numai dupa verificari experimentale si cu anumite riscuri.
5.4. PROCESUL DE DIFERENTIERE A ARBORILOR IN ARBORET
In cadrul unei populatii, ca si a speciei in general, variabilitatea genetica si fenotipica a organismelor este apreciabila. Practic, desi suficient de asemanator pentru a se incadra in specia respectiva, fiecare organism prezinta anumite particularitati care il individualizeaza. Acestea sunt de cele mai multe ori, de ordin cantitativ. Si la arborii crescuti in masiv, se constata aceste deosebiri, inca din primele faze de dezvoltare. Acestea se concretizeaza in diferentierea lor tot mai pronuntata odata cu inaintarea lor in varsta.
Diferentierea arborilor se manifesta in primul rand prin caracterele morfologice cantitative: inaltime, grosime, marimea coroanei, biomasa foliara, ca si prin intensitatea proceselor fiziologice: asimilatie, dezasimilatie, crestere, inflorire, fructificatie etc. Cauzele sunt unele variatii din genom, variatiile din mediul abiotic, dar si relatiile biocenotice.
La arbori, de exemplu, este importanta presiunea concurentiala provocata de starea de masiv.
Cea mai evidenta diferentiere a arborilor este cea pe inaltime, corelata insa si cu cea pe grosime si cea pe marimea si simetria coroanei. Aceasta diferentiere se produce din cauza tendintei de a capta cat mai multa lumina, intr-un masiv care limiteaza accesul la acest factor esential pentru viata plantelor. Arborii care au o predispozitie genetica, care au luat nastere dintr-o samanta mai mare care a germinat mai repede, intr-un macromediu mai umed sau mai bogat in ioni vor creste mai intens, isi vor dezvolta mai puternic coroana, vor ajunge deasupra altor arbori. Astfel, in masivul creat de coronamentul arboretului, se va produce o diferentiere a pozitiei coroanei arborilor, determinata de diferentele de inaltime. Unii arbori vor avea coroane mari, aproape in intregime deasupra masivului, altii vor avea coroane mai mici, incadrate jumatate sau mai mult in masiv, altii vor ramane chiar sub coroanele celorlalti arbori iar coroanele lor vor fi reduse ca marime.
Silvicultorul german Kraft, a evidentiat aceasta diferentiere printr-o clasificare pozitionala a arborilor pe 5 clase(figura 5.3.):
clasa I - arbori predominanti cei mai inalti, cu coroane mari, situate in buna parte deasupra masivului,
clasa II - arbori dominanti, cu inaltimi si coroane ceva mai mici, situate in masiv, dar cu jumatatea superioara luminata,
clasa III - arbori codominanti cu inaltimi si mai reduse si cu coroane mai mici, asimetrice, inghesuite intre cele ale arborilor dominanti,
clasa IV - arbori dominati, cu inaltimi mici, coroane foarte mici situate sub coroanele celorlalti arbori, numai cu varful in lumina,
clasa V - arbori deperisanti cu coroane aflate in intregime sub coroanele clorlalti arbori, inca vii dar slab dezvoltati (subclasa V a), sau uscati (subclasa V b).

Figura 5.3.
Diferentierea pe inaltime nu are insa caracter definitiv. Arborii, mereu in dezvoltare, pot sa acceada intr-o clasa superioara, dupa cum pot sa si decada in una inferioara. Aceasta dinamica este mai bine reflectata in clasificarea elaborata de IUFRO si care tine seama de inaltime, vigoarea de crestere si tendinta de modificare a pozitiei in arboret.
Clasificarea este zecimala si are urmatoarea forma:
Tabelul 5.5.
Clasificarea pozitionala a arborilor IUFRO
|
Criteriul |
|||||
|
Inaltime |
Cod |
Vigoare de crestere |
Cod |
Pozitie ierarhica |
Cod |
|
Arbori in plafonul superior |
Potential de crestere ridicat |
Tendinta de avansare | |||
|
Arbori in plafonul mijlociu |
Potential de crestere normal |
Tendinta stationara | |||
|
Arbori in plafonul inferior |
Potential de crestere redus |
Tendinta de regresie | |||
Daca un arbore are codul 123 inseamna ca este in plafonul superior, cu potential de crestere normal, dar cu tendinta de regresare ierarhica in plafonul inferior.
Diferentierea arborilor se produce in tinerete in faza denumita chiar dupa acest proces (intre 15-40 de ani in raport cu specia si statiunea). Dar procesul continua, cu intensitate mai slaba si la varste mai mari cand se produc schimbari pozitionale determinate mai ales de potentialul de crestere, schimbari in accesul la lumina prin disparitii de arbori vecini etc.
Pentru silvicultor cunoasterea diferentierii pozitionale a arborilor este baza de referinta pentru interventiile de conducere a arboretelor. De la inceput, pornind de la aceasta diferentiere, s-au format cele doua scoli de conducere a arboretelor:
scoala germana, care preconiza extragerea doar a arborilor dominati si deperisanti (cum se spunea figurat „ingroparea mortilor”)
scoala franceza, care preconiza interventia in toate plafoanele arboretului. Aceasta conceptie este si cea actuala.
5.5. PROCESUL DE CRESTERE SI DE ACUMULARE A LEMNULUI IN ARBORET
In arboret, din cauza masivului care conditioneaza o competitie accentuata pentru resursele nutritive, cresterea fiecarui arbore in parte este mai redusa, dar pe arboret, din cauza numarului mare de arbori, este totusi ridicata.
In arboret cresterea arborilor capata anumite particularitati. Astfel, cresterea in inaltime este mai mare decat la arbori izolati din cauza tendintei arborilor de a avea coroana in lumina. In masiv arborii vor avea deci trunchiuri mai inalte. Coroanele fiind insa mai mici si cantitatea totala de lemn produs mai redusa, cresterea in diametru nu va fi prea mare.
In arboret cresterea si acumularea de lemn este foarte diferita la arborii din diferite categorii pozitionale. Cele mai mari cresteri si acumulari de lemn se produc in arborii predominanti, cele mai mici in arborii dominati. Pe acest fapt se bazeaza scoala germana care preconiza ca prin lucrarile de conducere sa fie eliminati acesti arbori.
In arboretele pluriene, in care masivul se creaza in planuri diferite si cantitatea de lumina disponibila pentru arborii mai inalti este mai mare, de acest plus de lumina se folosesc mai ales arborii grosi si de grosime mijlocie care ajung la grosimi mai mari decat in arboretele echiene(figura 5.6.).
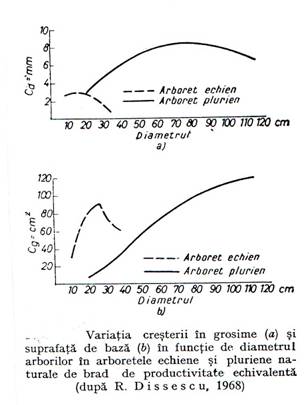
Figura 5.4.
Pentru a determina indicii dimensionali si productivitatea de lemn a arboretului denumit in silvicultura crestere, se executa doua inventarieri a arborilor din arboret la un anumit interval de timp (5 sau 10 ani), se calculeaza inaltimile si diametrele medii pe elemente de arboret, iar suprafata de baza si volumul si pe elemente de arboret si pe arboret in ansamblu.
Din diferentele ce se constata la dimensiuni si la volume intre cea de a doua si prima inventariere rezulta cresterile inregistrate in perioada respectiva.
In silvicultura se vorbeste de cresterea curenta produsa intr-un an si de crestere medie ce rezulta din impartirea cresterii pe un interval de timp la numarul de ani din acea perioada(figura 5.7.).
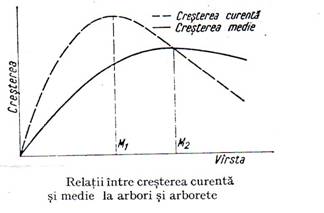
Figura 5.5.
Se vorbeste de asemenea de productia si productivitatea arboretului total cind se iau in considerare toti arborii, inclusiv cei ce s-au eliminat natural sau au fost extrasi de silvicultor prin lucrari de conducere si productia si productivitatea arboretului principal, cand se iau in considerare numai arborii ramasi dupa extrageri.
Pe baza inaltimii si diametrului mediu se poate determina arborele mediu pe fiecare element de arboret care are si suprafata de baza si volumul medii pe acel element.
5.6. PROCESUL DE ELAGARE NATURALA A TRUNCHIULUI ARBORILOR IN ARBORET
Starea de masiv care forteaza cresterea mai accentuata in inaltime a arborilor, determina si formarea de noi ramificatii pe trunchiul lor la inaltimi tot mai mari. Ramurile formate anterior, mai jos pe trunchi, sunt din ce in ce mai umbrite, iar in momentul cand lumina devine insuficienta pentru fotosinteza, frunzele de pe aceste ramuri si apoi ramurile se usuca si se desprind de trunchi. Locul de desprindere se cicatrizeaza prin acoperire cu tesuturile vii ale scoartei si apoi se acopera de cresterile ulterioare ale lemnului. Acesta este procesul de elagare al arborilor care confera trunchiului lor infatisarea speciala, curata de ramuri, care atesta cresterea in masiv. La arborii izolati elagarea nu se produce sau se elagheaza doar ramurile inferioare, umbrite de celelalte ramuri.
Inceperea si intensitatea procesului de elagare depinde de specie. La speciile de lumina incepe mai repede si este mai mare, la speciile de umbra intarzie si este mai redusa. Intensitatea depinde si de bonitatea stationala, fiind mai mare in statiuni de bonitate superioara, mai mica in cele cu bonitate inferioara.
Prin procesele de crestere in inaltime si de elagare coroana arborilor se deplaseaza mereu spre varf dar la speciile de lumina ramane putin dezvoltata pe inaltime, pe cand la cele de umbra capata extensiune mai mare. Acest lucru are consecinte asupra acumularii de lemn care depinde de cantitatea de frunze si deci, si de marimea coroanei.
Elagarea arborilor pe inaltimi cat mai mari ale trunchiului este de interes in silvicultura pentru a obtine cat mai mult lemn de calitate, fara sau cu putine noduri.
Dar, in acelasi timp, silvicultorul are interesul ca profunzimea coroanei si masa de frunzis sa fie suficienta pentru a intretine o acumulare ridicata de lemn. Problema care se pune este de a gasi solutia optima intre aceste doua tendinte opuse.
De regula, coroana trebuie sa cuprinda cel putin o treime din inaltimea arborelui.
5.7. MODIFICAREA FORMEI TRUNCHIULUI ARBORILOR IN ARBORET
Forma trunchiului arborilor este rezultatul combinat al cresterii in inaltime si grosime, destul de inegale, atat in timp cat si pe diferitele zone ale trunchiului.
Trunchiul arborilor are forma de paraboloid, diferit insa pe specii. In acest paraboloid sunt doua zone de crestere in grosime mai accentuate: la baza trunchiului si sub coroana, ca zone de rezistenta sporite. Ultima zona se deplaseaza in sus odata cu coroana, astfel ca forma trunchiului, in masiv, capata o cilindricitate din ce in ce mai pronuntata fata de arborele izolat, la care forma trunchiului este conica.
In general, prin cresteri compensatorii, in diferite zone ale trunchiului, solicitate mecanic din diferite cauze, exista tendinta permanenta de a crea forma trunchiului cea mai rezistenta la impacturi, care este cea
conic-cilindrica. In situatii de impact continuu, de exemplu de vant, forma trunchiului se adapteaza solicitarilor mecanice respective si poate fi ovalizata.
5.8. DINAMICA CICLICA IN ARBORET. SUCCESIUNEA ARBORETELOR
5.8.1. Dinamica ciclica a arboretului
Ca orice fenomen biologic, arboretul este supus la schimbari de diferite feluri avand o dinamica diurna, anuala, multianuala. Aceste dinamici au caracter ciclic pentru ca, desi sub influenta lor arboretul se schimba momentan, se produce o continua revenire a lui la compozitia si structura ce-i sunt proprii.
Exista insa si o dinamica succesionala care determina inlocuirea unui arboret prin altul cu compozitie si structura complet deosebite.
Dinamica ciclica diurna se refera la schimbarile ce se produc in arboret in decurs de 24 de ore. In anotimpurile in care arboretul este activ poate fi vorba despre desfacerea mugurilor, cresterea si miscarea frunzelor, desfacerea florilor, diseminarea, caderea frunzelor.
In cursul a 24 de ore se produc procesele fiziologice de fotosinteza (ziua), de respiratie si transpiratie (ziua si noaptea), de formare a lemnului etc.
Dinamica ciclica anuala se refera la schimbarile produse in cursul anului, in special cele fenologice, foarte usor de sesizat (inmugurire, infrunzire, inflorire, maturarea fructelor/semintelor, diseminarea, caderea frunzelor), cresterea in inaltime, grosime, volum.
In dinamica anuala poate interveni si o crestere sau scadere a populatiei sau populatiilor arboretului fie prin formare de noi indivizi, intr-un an cand se produce regenerarea naturala, fie prin uscarea altor indivizi.
Dinamica multianuala se refera la schimbarile care se produc in compozitia si structura arboretelor echiene cu varsta, de-a lungul ciclului lor de viata.
Se disting in acest sens acele faze ecologice mentionate in sectiunea 3.1.2. (faza de constituire a semintisului, faza de competitie a tineretului, faza de diferentiere a arborilor tineri, faza de maturitate, faza de imbatranire, faza de dezagregare).
Florescu I. (1981) arata ca in silvicultura se cunosc 4 etape de dezvoltare a arboretului:
etapa embrionara, de formare a semintei,
etapa tineretii, dureaza de la formarea plantulelor pana la inceperea fructificatiei,
etapa maturitatii, intre inceperea fructificatiei si inceperea depericiunii arborilor,
etapa batranetii cand arborii batrani dispar.
In silvicultura se foloseste o alta impartire bazata pe criterii mixte, de dezvoltare si dimensionale.
Fazele „silviculturale” de dezvoltare a arboretului in raport cu varsta sunt:
faza de semintis, de la aparitia in masa a puietilor pana la formarea starii de masiv,
faza de desis, pana la inceperea elagajului natural si pana la inaltimea arborilor de 2-2,5 m,
faza de nuielis, de la inceperea elagajului natural pana cand arborii au grosimi de 5 cm,
faza de prajinis, cu arbori avand grosimi de 5-(8)10 cm,
faza de paris, cu arbori avand grosimi de (8)10-18(20) cm,
faza de codrisor, cu arbori avand grosimi intre 21-35 cm,
faza de codru mijlociu, cu arbori avand grosimi peste 35 cm,
faza de codru batran, cu arbori lancezi si in uscare,
Corespondenta intre fazele ecologice, etapele de dezvoltare si fazele „silviculturale” in dinamica multianuala a arboretului sunt date in tabelul 5.6.
Tabelul 5.6.
|
Fazele ecologice (dupa N. Donita) |
Etapele de dezvoltare (dupa I. Florescu) |
Fazele „silviculturale” (dupa tratatele clasice) |
|
Etapa embrionara | ||
|
Faza de constituire a semintisului |
Etapa tineretii |
Faza de semintis |
|
Faza de competitie a tineretului |
Faza de desis Faza de nuielis |
|
|
Faza de diferentiere a arborilor tineri |
Faza de prajinis Faza de paris |
|
|
Faza de maturitate |
Etapa maturitatii |
Faza de codrisor Faza de codru mijlociu |
|
Faza de imbatranire |
||
|
Faza de dezagregare |
Etapa batranetii |
Faza de codru batran |
Fazele silviculturale sunt importante pentru ca de ele sunt legate interventiile silvotehnice de ingrijire si conducere a arboretului.
In arboretele pluriene aceste faze sunt, de asemenea prezente dar pe suprafete reduse. Pe intregul arboret ele coexista. De aceea, in asemenea arborete toate masurile de ingrijire si conducere se fac simultan si nu decalat in timp ca in arboretele echiene.
5.8.2. Succesiunea arboretelor
Este un proces obisnuit atunci cand se schimba radical statiunea (atat ca geotop cat si ca ecotop).
Speciile vechiului arboret, nemai avand adaptarile necesare pentru a se putea dezvolta in noile conditii, dispar si in locul lor patrund specii care au aceste adaptari.
Succesiunile pot fi rapide sau lente. Un exemplu de succesiune rapida este cea de inlocuire a unui arboret de mesteacan, care a ocupat primul o suprafata fara arboret, sub care s-a instalat apoi un semintis de molid. Acesta, la varste mai mari, va elimina mestecanisul, rezultand un arboret de molid.
Aici este vorba de relatii de competitie intre specii si de rapiditatea ocuparii teritoriului.
Dar succesiunile pot fi si lente. Cel mai bun exemplu il constituie modificarile de compozitie a arboretelor in padurile ce s-au format in postglaciar, pe teritoriul tarii. In rastimp de circa 12000 ani, in raport cu schimbarea climatului, s-au succedat biocenoze edificate de arborete de pin, de molid, de stejar, apoi de arborete de carpen, de fag, si in ultima perioada, si de arborete mixte de fag si brad.
Pe silvicultor il intereseaza in special succesiunile nedorite, de inlocuire a arboretelor din specii valoroase prin cele de specii mai putin valoroase. Asa este, de exemplu, inlocuirea stejaretelor prin carpinete.
Dar, in ultimul timp, din cauza schimbarilor climatice majore, provocate de efectul de sera, silvicultorii se vor confrunta si cu succesiuni de tipul lent, dar probabil mult grabite, care vor duce la schimbarea compozitiei arboretelor din biocenozele forestiere actuale intr-o directie inca putin cunoscuta.
5.9. RELATIILE ARBORETULUI CU ALTE COMPONENTE BIOCENOTICE
Relatii cu alte straturi ale biocenozei. Desi straturile arbustilor si ierburilor au pozitie subordonata fata de arboret, acestea au si influente asupra acestuia. Un strat de arbusti dezvoltat, mai ales in cazul arboretelor cu grad de acoperire mai redus, impiedica intelenirea solului si pierderi de apa, creand conditii de lumina nefavorabile pentru ierburi. Multe pasarele insectivore cuibaresc in arbusti. Arbustii si ierburile maresc biodiversitatea biocenozei, ceea ce mareste stabilitatea acesteia, ca si a arboretului. In schimb un strat al ierburilor, ca si a arbustilor puternic dezvoltate pot impiedica regenerarea naturala prin concurenta pentru lumina, apa, ioni din sol.
Relatii cu populatiile consumatoare. Fiind principalii producatori din biocenoza, de arbori se leaga foarte multe lanturi trofice. Unele populatii consumatoare, de ordinul I, in caz de inmultire in masa, pot afecta grav arboretul prin defoliere, prin consumul liberului, al lemnului, al semintelor, puietilor. In schimb consumatorii de ordinul II sunt favorabili arboretului reducand populatiile fitofage. Silvicultorul are tot interesul sa le mentina si sa le foloseasca (pasarele insectivore, furnici, bufnite etc.)
Relatii cu populatiile descompunatoare sunt indirecte.
Prin necromasa furnizata de arboret se asigura viata a numeroase populatii de ciuperci si microfauna. Aceste populatii, prin descompunerea materialului mort si prin participare la formarea humusului si la evolutia solului, sunt utile arboretului, pentru ca asigura autofertilizarea si evolutia solului si deci, mentinerea productivitatii arboretului.
Relatiile cu vanatul ierbivor poate avea efecte negative in cazul depasirii efectivelor optime, adica suportabile din punct de vedere biocenotic.
Vanatul prea numeros distruge in masa semintele (porcii mistreti), puietii (cerbul, capriorul) dar si arborii tineri prin smulgerea scoartei sau ranirea arborilor prin frecarea coarnelor.
Pentru a interveni activ in relatiile arboretului cu alte componente biocenotice silvicultorul trebuie sa cunoasca bine structura trofica a biocenozei, si toate relatiile care pot afecta dar, care pot si favoriza arboretul.
5.10. RELATIILE ARBORETULUI CU STATIUNEA CA GEOTOP SI CA ECOTOP
Aceste relatii sunt foarte stranse pentru ca arboretul isi extrage energia si materia cu care produce biomasa din statiune, dar si influenteaza puternic statiunea.
Statiunea prin caracteristicile ei climatice si edafice are, asa cum s-a aratat, rol selectiv si de control in formarea si fiintarea arboretului. Prin cantitatea de energie si materie pe care o furnizeaza (bonitatea stationala) statiunea determina productivitatea arboretului, iar prin unele conditii stationale (relief, roca, sol) si stabilitatea arboretului (soluri profunde pe roci friabile, forme de relief protectoare de vant etc.)
Influenta modificatoare a biocenozei asupra statiunii s-a tratat mai pe larg in capitolul 3 (3.1.4.). Este doar de retinut ca influenta modificatoare a biocenozei forestiere asupra statiunii se datoreste in cea mai mare parte efectului generator si protector de mediu al arboretului.
De aceea, in asigurarea serviciilor de mediu pe care le ofera padurea, silvicultorul trebuie sa se preocupe, in primul rand, de structura arboretului, de realizarea structurii optime pentru realizarea fiecarui serviciu.
5.11. RELATIILE ARBORETULUI CU STRATUL DE NECROMASA
Necromasa depusa pe sol si aflata in diverse stadii de descompunere are efecte benefice, dar si nefavorabile asupra arboretului.
Efectele benefice se concretizeaza in acoperirea solului care favorizeaza infiltratia, impiedica eroziunea, pierderile de apa prin evaporatie si pierderile de caldura, precum si in fertilizarea solului prin formarea humusului si mineralizarea materiei organice.
Efecte nefavorabile, mai ales asupra regenerarii arborilor, se concretizeaza in formarea de acumulari de litiera proaspata sau de litiera slab descompusa (pe soluri foarte acide) care impiedica regenerarea naturala, ca si incalzirea solului in climate reci.
Pastrarea necromasei pe sol in padure este obligatorie, dar in cazul acumularii prea mari se impun masuri de activizare a descompunerii (prin rarirea arboretului care sa permita patrunderea de mai multa caldura si apa la
|
