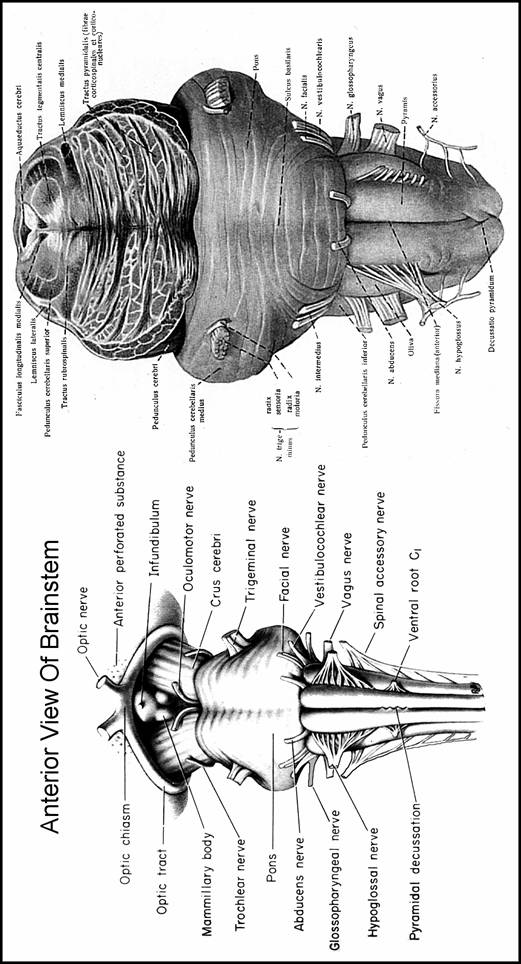
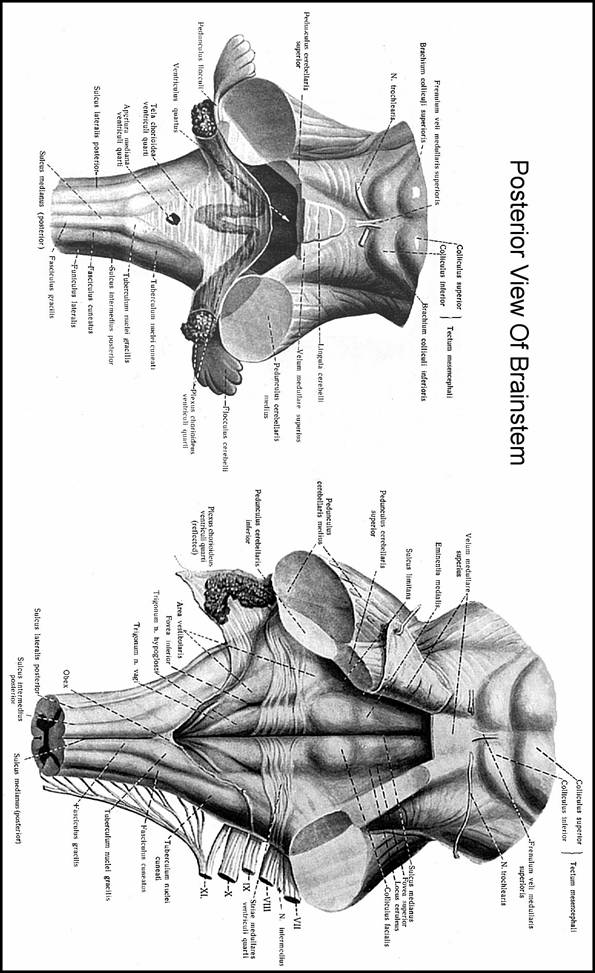
BRAIN STEM
It represents that segment of CNS, which is interposed between spinal cord cerebrum and cerebellum. It has an oblique antero-superior position and consists of 3 components which, from inf. to sup. direction are: medulla oblongata (or spinal bulb pons Varolii and midbrain (or mesencephalon
MEDULLA OBLONGATA
It represents the smallest and the most caudal portion of the brain stem;
Limits
sup. limit (between medulla and pons) is represented:
anteriorly by the bulbo-pontine sulcus (or ponto-medullary sulcus) which has a transverse direction,
posteriorly by the striae medullares of the IVth ventricle which are some transverse thin bundles of white matter on the floor of the IVth ventricle
inf. limit (between spinal cord and medulla) is represented by an imaginary transverse plan that passes:
immediately under the pyramidal decussation or
above the first pair of cervical spinal nerves or
at the same level with foramen occipitale magnum.
General shape is of a pyramidal trunk that presents:
a large superior extremity continuous with the pons and
a narrow inferior extremity continuous with the spine;
External aspect - its surface presents all the sulci and fissures of the cervical spinal cord:
Anterior-median fissure continues rostrally the same fissure of the spine, and it ends rostrally at the lower margin of the pons (the bulbo-pontine sulcus) in a small triangular fossa called foramen caecum; it separates the 2 medullary pyramids and it is interrupted in the inf. part of medulla by the pyramidal decussation;
Posterior-median sulcus continues rostrally the same sulcus of the spine and it exists as long as it separates the 2 gracilis fascicles; this happens in the so-called closed part of the medulla which represents the inf. half of this segment; it finishes rostrally at the place where the 2 gracilis fascicles diverge and the central canal of the spine expands into the cavity of the IVth ventricle;
2 anterior-lateral sulci - they represent the rostral continuations of the same sulci of the spine; they separate the anterior face of medulla (the 2 pyramids) from its 2 lateral faces; their superior halves lie anteriorly to the olives of the medulla; for this reason they are also called the preolivary sulci and here the filets of hypoglossal nerve 353j99d (CN-XII) emerge at the medullary surface (they are somehow the equivalents of the anterior root of the spinal nerves);
2 posterior-lateral sulci - they represent the rostral continuations of the same sulci of the spine and they separate the 2 lateral faces of the medulla from its posterior face;
2 posterior-intermediate sulci - they represent the rostral continuations of the same sulci of the spine; they also separate the 2 fasciculi gracilis and cuneatus on the same side and diverge together with them when the IVth ventricle appears as a cavity;
Medullary surface also presents regions (faces) separated by the described sulci; these regions represent the continuation of the white columns of the spine (there are 3 regions on each side):
Anterior region - it is also called the medullary pyramid and lies between the anterior median fissure and the anterior-lateral sulcus of the same side; the pyramid has the aspect of an elongated superficial elevation that continues rostrally the corresponding anterior white column of the spine; its upper end is slightly constricted and here, between medulla and pons, emerges the root of abducens nerve (CN-VI); the pyramid contains the fibres of cortico-spinal tract which are going to form the pyramidal decussation;
Posterior region - it lies between the anterior and posterior lateral sulci; it presents:
the medullary olive - it is an oval prominence which is located in the anterior part of the superior half of this region; the olivary prominence is caused by the underlying principal inferior olivary nucleus; it is separated from the surface of this region by 2 sulci:
preolivary sulcus (see the anterior-lateral sulci) and
retroolivary sulcus (it lies posteriorly to the olive); from retroolivary sulcus emerge the rootlets of glossopharyngeal nerve (CN-IX) and vagus nerve (CN-X);
Some authors describe the emergences of CN-IX and CN-X at the level of the lateral region and not quite from retroolivary sulcus;
the rootlets of accessory nerve (CN-XI) emerge caudally to those of vagus nerve, at the level of the lateral region; these rootlets are numerous and they also emerge from the lateral funiculus of the superior cervical spinal cord; after their emergence, these rootlets join to form a single nervous trunk, which ascends and enters the cranial cavity through foramen magnum;
Posterior region lies between the 2 posterior-lateral sulci and it presents
a caudal half (the "closed part of the medulla") which continues the posterior funiculi of the spine; it has the same characteristics as the posterior faces of the cervical spine;
a cranial half - here appears the inferior triangle of the IVth ventricle which determine the 2 fasciculi gracilis to diverge; each fasciculus gracilis and cuneatus presents in its terminal part (immediately above their diverging place) a swelling called tuberculus gracilis (or clava) and tuberculus cuneatus, each one being determined by the corresponding underlying nuclei gracilis and cuneatus; cuneatus tuberculi have a higher localisation than gracilis ones; the 2 fasciculi gracilis and cuneatus seem to be continued rostrally by the inf. cerebellar peduncles, but in reality, fasciculi gracilis and cuneatus finish their fibres in the nuclei with the same name; the inf. cerebellar peduncles lie on the both inf. margins of the IVth ventricle and enter the white matter of cerebellum;
The rest of the post. region (the part between the 2 inferior cerebellar peduncles) is formed by the floor of the IVth ventricle (especially its inferior triangle) and for this reason it will be studied together with the other components of the IVth ventricle;
PONS VAROLII
It represents the intermediate segment of the brainstem, lying between medulla and midbrain.
Localisation
it lies - under the midbrain (caudally to the midbrain), above the medulla (rostrally to the medulla) and ventrally to the cerebellum.
Limits
The inferior limit (between medulla and pons) was described as the sup. limit of medulla.
The superior limit (between pons and midbrain) is represented by the ponto-peduncular sulcus which surrounds the anterior and lateral faces of the brainstem at this level;
External aspect
it presents 2 faces
Ventral face:
its aspect: is prominent and its convexity is greater transversally and lesser vertically; it is sustained by the dorsal face of sphenoid bone (dorsum sellae) and by the basillary part of occipital bone (these bony parts form a large sulcus called clivus where medulla and pons are lying);
it consists of transverse fibres that - pass like a bridge over the midline (that's why it is called "pons"); these fibres converge on each side in order to form the 2 middle cerebellar peduncles
its surface presents:
sulcus basilaris that contains the basilar artery - it is a superficial median vertical sulcus which is bounded bilaterally by 2 rounded eminences caused by the cortico-spinal fibres which descend in the pons;
apparent origin of CN-V - it lies laterally to the 2 eminences caused by corticospinal fibres and at the transverse midleve1 of the pons; it has a thin motor root (it lies superomedially) and a thick sensory root (it lies inferolaterally)
The vertical line through this apparent origin of CN-V represents the conventional limit between the ant. face of pons and the middle cerebellar peduncles;
on the ventral face we can also describe the apparent origins of CN-VII, CN-VII bis and CN-VIII lying at the level of bulbo-pontine sulcus; above the olive lie CN-VII, and VII-bis) and near the inferior cerebellar peduncle lies CN-VIII;
Dorsa1 face cannot be seen due to the cerebellum that covers it; it forms the sup. triangle of the floor of the IVth ventricle and a part of its roof (sup. medullary vellum); for this reason it will be studied together with the other components of the IVth ventricle;
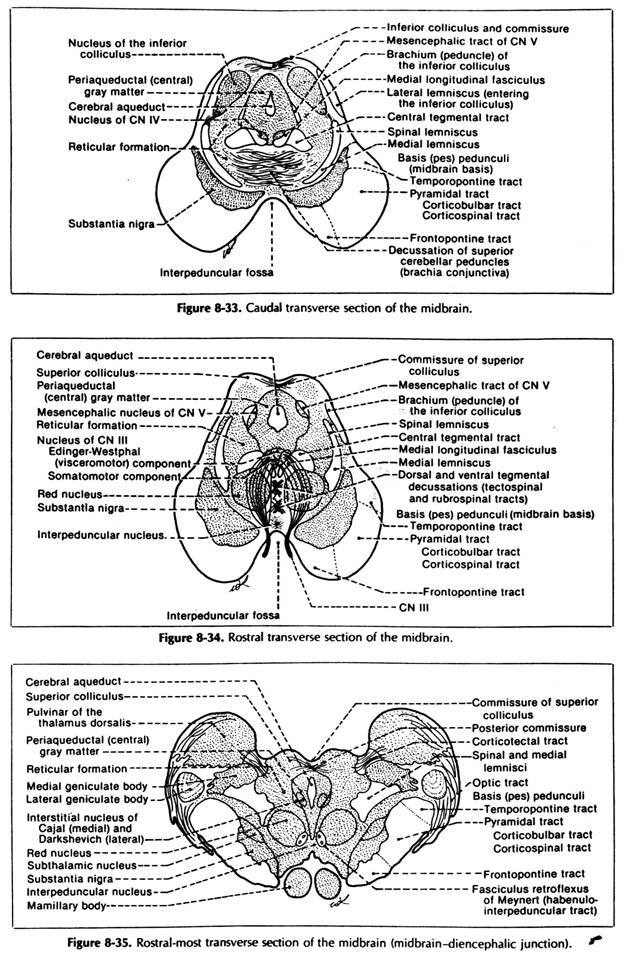
MIDBRAIN (MESENCEPHALON)
It represents the shortest and most rostral part of the brainstem.
Limits
inf. limit (between pons and midbrain) is represented by ponto-peduncular sulcus;
sup. limit (between midbrain and diencephalon) is represented by a horizontal imaginary plan that passes immediately above the optic chiasma and optic tracts;
Course and relations: it traverses the hiatus in the tentorium cerebelli which is represented by the tentorial notch; laterally to the midbrain lie the 2 parahippocampal gyri which hide its lateral sides; its long axis is oriented antero-superiorly; for an easier description, the anterior part of midbrain (its base) is divided into 2 halves (a right and a left one) called cerebral peduncles
it presents 2 faces
ventral face presents:
the 2 crurae cerebri - are 2 big cylindrical elevations diverging in sup. direction after they emerge from the pons; -they enter the infero-medial part of each cerebral hemisphere; they represent the "base" of the midbrain and consist of white matter only, which is separated from the tegmentum by substantia nigra;
interpeduncular fossa - it is a triangular depression between the 2 diverging crurae cerebri; it contains:
posterior perforated substance - it represents the caudal median part of the area that belongs to the interpeduncular fossa; it presents many small foramina where the branches of post. cerebral arteries pass inside nervous substance;
medial sulcus - it is a paired-sulcus, each one lying on the medial face of each crus cerebri; it presents the apparent origin of the oculomotor n.
dorsal face - it presents:
the 4 quadrigeminal colliculi - they are 4 rounded eminences (2 sup. ones and 2 inferior ones); they lie under the pineal gland and post. white commissure; above the sup. medullary vellum they are partially covered on each side by the pulvinar of the thalamus (its dorsal part); they are separated by a cruciform sulcus
frenulum of the sup. medullary vellum - it is a small fold of white matter that prolongs down upon the sup. medullary vellum beginning from the vertical sulcus between the 2 inferior quadrigeminal colliculi;
apparent origin of trochlear n.- it is located immediately lateral to the frenulum of the sup. medullary vellum; this nerve surrounds the lateral aspect of cerebral peduncle on its side and reaches anteriorly in the ponto-peduncular sulcus;
2 brachia of the superior quadrigeminal colliculi - they are small elongated prominences (2 in number) ascending ventro-laterally from the latera1 aspect of each sup. quadrigeminal colliculus; it passes under the pulvinar and continues partially in the optic tract and partially in the lateral geniculate body on the same side;
2 brachia of the inferior quadrigeminal colliculi - they are small elongated prominences (2 in number) ascending ventrally from each lateral aspect of the each inf. quadrigeminal colliculus;
the ventral and dorsal faces of the midbrain meet at their lateral extremities and here they are separated by a longitudinal lateral sulcus which is somehow parallel with the longitudinal axe of the corresponding cerebral peduncle;
FOURTH VENTRICLE
It is a cavity of brainstem that has a complex shape and lies:
ventrally to the cerebellum and
dorsally to the pons and to the sup. 1/2 of the medulla (which is called the "open part of the medulla");
It has the general shape of a diamond. It has the next components
a floor (ant. wall) called "rhomboid fossa" due to its rhomboidal shape
a roof (post. wall)
4 margins
2 sup. margins formed by the sup. cerebellar peduncles,
2 inf. margins formed by gracilis and cuneatus tubercles and by inferior cerebellar peduncles
4 angles
a sup. angle - it is formed by the 2 sup. margins; at its level the fourth ventricle is continued with the cerebral aqueduct Sylvian aqueduct
an inferior angle - it is formed by the 2 inf. margins of the ventricle; at its level the fourth ventricle is continued with the central canal of the "closed part" of the medulla (the inf. 1/2 of medulla);
2 lateral angles - they are formed by the sup. and inf. Margins; they form the lateral recesses of the fourth ventricle.
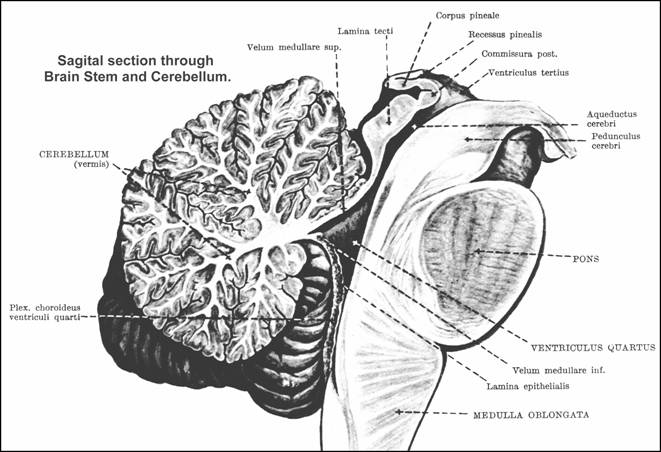
The roof - from sup. to inf. direction it is formed by:
a part of the sup. cerebellar peduncles,
sup. medullary vellum
inf. medullary vellum
tella choroidea of the fourth ventricle (tella choroidea inferior)
ligula (taenia of the fourth ventricle)
obex
We shall describe each of these components:
Superior cerebellar peduncles - they are 2 bundles of fibres emerging from the white matter of cerebellum; they run in antero-superior direction and converge as they approach the inf. quadrigeminal colliculi; their medial parts form the 2 sup. margins of the fourth ventricle and at the same time they form a part of its roof (especially at the place where they converge superiorly)
Superior medullary vellum - it is a thin lamina of white matter lying in the angular space between the 2 sup. cerebellar peduncles; it is continued dorsally with the white core of the cerebellum; its dorsal face is covered by lingula cerebelli (the sup. extremity of the vermis);
Inferior medullary vellum - this vellum is a thin and narrow transverse lamina of white matter which forms the roof of the fourth ventricle between its lateral angles; it has quadrangular shape; it presents:
a sup. margin - it is convex and continues with the white matter of cerebellum;
an inf. margin - it is concave; apparently it is free, but it continues inferiorly with tella choroidea of the fourth ventricle;
2 very short lateral margins which are continued with the white matter of the inf. cerebellar peduncles;
*** Fastigium - it represents the angle of the roof, which is formed between sup. and inf. medullary vella (this angle is opened anteriorly)
Tella choroidea inferior
it forms the roof of the inf. triangle of the fourth ventricle bellow inf. medullary vellum; it is not formed by nervous matter; it consists of 2 layers:
ependyma - it is the internal layer and it represents the mesothelium that covers the internal surface of all the cavities of CNS,
pia mater - it forms the external layer
superiorly tella choroidea is reflected on the inf. surface of cerebellum (in fact, here, by reflection, pia mater forms 2 layers which are more or less adherent between them)
inferolaterally tella becomes inserted on the inferior margins of the fourth ventricle (the place of insertion is marked by the taenia of the fourth ventricle);
between the 2 layers of tella choroidea lies the choroid plexus of the fourth ventricle; this plexus is formed by cells of ependyma which are modified and become secretory epithelium having a rich blood supply; choroid plexus lies inside an invagination of the ependyma and thus it becomes projected inside the cavity of the fourth ventricle; it has the general shape of the letter "T" having 2 vertica1 limbs which are joined inferiorly; the lateral extremities of the horizontal limb enter the lateral foramina (Luschka) and reach the subarachnoidian space;
tella choroidea presents 3 orifices which represent the only communication between the cavities of CNS and subarachnoidian space; these orifices are:
1 foramen of Magendie - it is a single foramen lying on the median line above the choroid plexus of the fourth ventricle;
2 foramina of Luschka (or foramina of Key and Retzius), each one lying at the level of tella choroidea above the lateral recesses of the fourth ventricle; through these foramina, the lateral extremities of the horizontal limb of the choroid plexus protrude into the subarachnoidian space;
ligula - it is also called "taenia of the fourth ventricle"; it consists of 2 narrow bands of white matter lying at the place where tella choroidea inferior inserts on the 2 inferior margins of the fourth ventricle; first, the 2 taeniae lie on the medial margins of the 2 fasciculi gracilis at the place where they diverge and then they run antero-superiorly following the inferior margins of the fourth ventricle; finally they turn laterally over the restiform bodies (inf. cerebellar peduncles) and reach as latera1 as the lateral recesses; the medial borders of the 2 taeniae are continued with the tella choroidea of the fourth ventricle;
Obex - it is a thin triangular lamina of grey matter that covers the inferior angle of the cavity of the fourth ventricle; it represents the superior extremity of the grey matter which forms the grey commissure of the spina1 cord; it is covered by ependyma both on its anterior and posterior surfaces;
The floor
It is also called "rhomboid fossa" due to its rhomboidal shape.
It is formed by:
the dorsal surface of the pons
the dorsal surface of the sup. 1/2 of the medulla (the "opened part" of medulla).
It consists of 2 triangles
sup. triangle (it belongs to the pons) and
inf. triangle (it belongs to the medulla). These 2 triangles are separated by striae medullares that arise from the median longitudinal sulcus and run in lateral direction to enter the inf. cerebellar peduncles
It is traversed by the median longitudinal sulcus, which runs vertically between the sup. and inf. angles of the fourth ventricle;
it is also traversed by striae medullares which run horizontally between the 2 lateral angles (or recesses) of the fourth ventricle; these are thin transverse bundles of white matter that emerge from medial longitudinal sulcus and run to the inf. cerebellar peduncles where they enter;
Superior triangle - it presents:
2 sulci limitans - are 2 parallel sulci that lie on both sides of median longitudinal sulcus; its sup. part forms itself the lat. limit of the floor - it widens superiorly into an angular depression called fovea superior; it presents in its sup. part a bluish-grey locus coeruleus that covers a group of pigmented neurons called nucleus coeruleus; this nucleus is considered as a part of the pontine reticular formation, has neuromelanin in some of its neurons and represents the largest group of noradrenergic neurons in mammalian brains;
2 eminentia teres - they lie between each sulcus limitans and median longitudinal sulcus; they are spindle-shaped;
2 facial colliculi - they are 2 more prominent rounded formations in the middle of eminentia teres; they are determined by the fibres of facial nerve which surround the nucleus of abducens nerve located under the surface of these colliculi;
conductor sonorius - is a bundle of fibres which is connected in its inf. part with striae medullares and it ascends laterally crossing sometimes fovea superior
area vestibularis - is a paired area lying on both sides of the eminentia teres; it extends in the lateral recess where it becomes the auditory tubercle and in the inferior triangle where it forms the most lateral trigone; under these areas lie the vestibular nuclei;
Inferior triangle - it presents:
fovea inferior - it is an angular sulcus with the apex at the level of striae medullares of the same side and 2 limbs (a medial and a lateral one) diverging inferiorly; it borders:
a triangular area on each side of median longitudinal sulcus which is called the vagal trigone; under this trigones lie the 2 dorsal vagal nuclei;
hypoglossal trigone - it lies between the medial limb of each fovea inferior and the median longitudinal sulcus; under this trigone lies the nucleus of hypoglossal n.
area vestibularis -this is the prolongation of the same area of the sup. triangle of the floor of the fourth ventricle; it lies laterally to the lateral limb of fovea inferior;
funiculus separans - it is a narrow translucent fold which crosses the inf. part of the vagal trigone; it is covered by thickened ependyma containing tanycytes
area postrema - it lies between funiculus separans and tuberculum gracilis - it is considered as a circumventricular organ (it is the only paired organ of this type) and it is formed by a specialized ependyma (ependymocytes and tanycytes) which are involved in:
secretion of CSF,
transport of neurochemicals from CSF to the subjacent structures (neurons, glial cells or vessels) and also in opposite direction,
chemoreception (especially area postrema)
INTERNAL STRUCTURE OF BRAINSTEM (General rules)
All the 3 segments of the brainstem (medulla, pons and midbrain) have a similar plan of internal organisation, which is based on the concept of tectum tegmentum and basis. These are 3 continuous laminae of nervous substance that extend longitudinally on the entire length of the brainstem.
Tectum
it is the plate of nervous tissue of the brainstem which lies dorsally to the transverse plan that passes along the Sylvian aqueduct ("tectum"= roof);
it consists of:
quadrigeminal plate at the level of the midbrain,
sup. medullary vellum at the level of the pons,
inf. medullary vellum at the level of the medulla.
tectum covers the next cavities of the CNS:
Sylvian aqueduct cerebral aqueduct) at the level of the midbrain,
fourth ventricle at the level of the pons and medulla.
Sup. and inf. medullary vella form an angle at the level of the roof of the fourth ventricle; this angle is called fastigium and is opened anteriorly; this angle is the peak (apex) of the cavity of the fourth ventricle;
in its structure tectum contains no
cranial nerve nuclei,
reticular formation,
long motor or sensory pathways which pass longitudinally through the brainstem.
Basis
it is the ventral part of the brainstem which is sustained by clivus;
composition:
descending cortical motor tracts
pyramidal tracts:
corticospinal tract,
corticobulbar tract
corticopontine tracts which are a part of cortico-ponto-cerebellar pathway
nuclei of the base of pons which are scattered inside the white substance at the level of the pontine basis.
differences in the composition of the bases of midbrain, pons and medulla:
Midbrain basis
it contains all 3 types of fibres:
corticospinal (crurae cerebri),
corticobulbar,
corticopontine;
it has no nuclei
Pontine basis
it is the largest one (it contains both tracts and nuclei),
the corticopontine fibres are finishing in the nuclei of the pontine basis where they synapse,
fibres which arise from these nuclei decussate, converge in order to form the middle cerebellar peduncles and finally they project to the cerebellar cortex;
the great majority of corticobulbar fibres enter the tegmentum at the level of midbrain and pons
all corticospinal fibres and some remaining corticobulbar fibres pass through the pontine basis and reach the medullary basis;
Medullary basis (medullary pyramids) - it is the smallest one because it contains:
just 2 small nuclei (arcuated nuclei which are the equivalent of the nuclei of the pontine basis),
a lesser number of fibres than in the other basal laminae of the midbrain:
a few corticobulbar fibres which synapse into the arcuated nuclei in the same way as the corticopontine fibres in the pontine nuclei; fibres which arise from arcuated nuclei are called anterior external arcuated fibres and they surround the external surface of medulla to enter the inf. cerebellar peduncles;
the corticospinal fibres;
The corticospinal fibres in the medullary basis form the medullary pyramids; their name comes from their pyramidal shape on transverse section and from their origin in the pyramidal neurons (Betz) of the cerebral cortex;
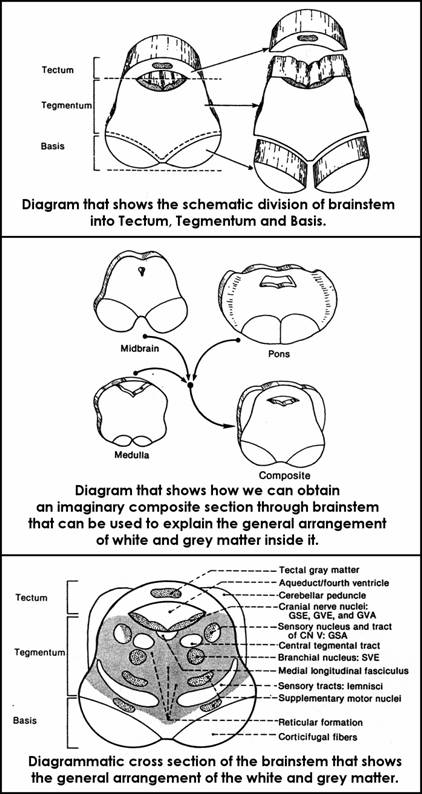
Tegmentum
it is the lamina of nervous tissue that lies between tectum and basis ("tegmen" = covering; here it means "covering of the basis");
the limit between tegmentum and basis at the level of the midbrain is made by substantia nigra;
composition:
it is the complicated part of the brainstem because it contains both grey and white matter
it consists of:
grey matter
motor and sensory nuclei of the cranial nerves III-XII (except CN-XI),
reticular formation,
proper nuclei.
white matter:
all the ascending sensory tracts that run from the spinal cord and cranial nerve nuclei to the cerebellum, thalamus or to the nuclei of the brainstem,
medial longitudinal fasciculus,
central tegmental tract which interconnects tegmentum, diencephalon, and basal nuclei,
cerebellar afferent and efferent fibres,
deep unnamed pathways in the reticular formation
scheme of the general composition of tegmentum
general somatomotor nuclei of CN and general sensory nuclei of CN - they lie in the dorsal part of tegmentum;
special visceral efferent (SVE) nuclei 1ie ventrolaterally to the general somatic efferent (GSE) nuclei;
relay motor nuclei proper nuclei of the brainstem) lie:
in the ventral part of tegmentum or
in the base of the pons;
These nuclei are: inferior olivary nuclei (in medulla), nuclei of the basis of the pons (in pons), red nucleus and substantia nigra (in midbrain);
the lemnisci (sensory tracts to the thalamus) run in the ventral and lateral parts of the tegmentum;
reticular formation fills in the space which is unoccupied by all the other structures;
Functional types of neurons and nervous fibres (or pathways):
Spinal nerves contain 4 functional components
GSE = general somatic efferent (they provide the motor innervation for striated muscles);
GVE = general visceral efferent (they provide motor innervation for smooth muscles and glands;
GSA = general somatic afferent (they represent the sensory innervation that comes from exteroceptors and proprioceptors)
GVA = general visceral afferent (they represent the sensory innervation that comes from interoceptors);
Cranial nerves contain all these 4 functional components (GSE, GVE, GSA, GVA) and more other 3 ones:
SVE = special visceral efferent (they represent the motor neurons and fibres that innervate the striated muscles derived from embryonic branchial arches)
SVA = special visceral afferent (they represent the sensory innervation of the special sensory receptors of endodermal origin - olphactory mucosa and taste buds
SSA = special somatic afferent (they represent the sensory innervation of the organs of special sense of ectodermal origin - the eye and the ear
|
Cranial Nerve (name and number) |
Functional type of its fibres |
Nucleus of Cranial Nerve and its localisation |
Peripheral target of Cranial Nerve Fibres (receptors and effectors) |
|
Olphactory n. CN-I- |
SVA |
- Special sensory fibres which innervate the receptors of the olphactory mucosa |
|
|
Optic n. CN-II- |
SSA |
- Special sensory fibres which innervate the rode and cone cells from the retina |
|
|
Oculomotor n. CN-III- |
GSE GVE |
Oculomotor nucleus in midbrain Edinger-Westphal nucleus in midbrain |
- Somatic motor innervation for the next extraocular mm.: sup. rectus, inf. rectus, medial rectus and inf. oblique; also for levator palpebrae sup. m. - Parasympathetic innervation of the eye. Preganglionic fibres synapse in the ciliary ggl. Postganglionic fibres innervate the circular mm. of the iris and the ciliary body. |
|
Trochlear n. CN-IV- |
GSE |
Trochlear nucleus in midbrain |
- Somatic motor innervation for the sup. oblique m. (extraocular m.) |
|
Abducent n. CN-VI- |
GSE |
Abducent nucleus in pons |
- Somatic motor innervation for the lat. rectus m. (the last extraocular m.) |
|
Trigeminal n. CN-V- |
SVE GSA |
Motor nucleus of CN-V- in pons Spinal nucleus of CN-V- in medulla oblongata Main sensory nucleus of CN-V- in pons and mesencephalic nucleus Of CN-V- in midbrain |
- Branchiomotor (somatic) innervation for the muscles of mastication, tensor tympani m. and tensor velli palatini m. - Somatic sensory innervation of the exteroceptors of the face, cornea, bucca1 mucosa and the ant. 2/3 of the tongue (not from the taste buds) - Proprioceptive sensory innervation (it is also somatic) from the muscles of mastication. |
|
Facial n. CN-VII- |
SVE GVE GVA SVA GSA |
Facial motor nucleus in pons Sup. salivatory nucleus in pons Nucleus of the solitary tract Nucleus of the solitary tract Spinal nucleus of CN-V- |
- Branchiomotor (somatic) innervation for the muscles of facial expression - Parasympathetic motor innervation for different glands. Preganglionic fibres synapse in the pterygopalatine ggl. (sphenopalatine ggl.). Postganglionic fibres innervate the lachrymal, nasal, and palatine glands. )Some authors describe the origin of these preganglionic fibres in the lachrymo-muco-nasal nucleus in pons (or it is also called lachrymal nucleus). Other preganglionic fibres (after some authors these originate exclusively in the sup. salivatory nucleus) will synapse in the submandibular ggl. Postganglionic fibres innervate the sublingual and submandibular salivatory glands. - Visceral sensory innervation of the soft palate and nasal cavity. - Sensory innervation of the taste buds in the ant.2/3 of the tongue. - Somatic sensory innervation of the external auditory meatus and external ear. |
|
Glossopharyngeal n. CN-IX- |
SVE GVE GVA SVA GSA |
Nucleus ambiguus Inf. salivatory nucleus Nucleus of solitary tract Nucleus of solitary tract Spinal nucleus of CN-V- |
- Branchiomotor innervation of the stylopharyngeal m. - Parasympathetic innervation of the parotid gland. Preganglionic fibres synapse in the otic ggl. Postganglionic fibres innervate the parotid gland. - Visceral sensory innervation of the pharyngeal mucosa and mucosa of the post.1/3 of the tongue. - Sensory innervation of the taste buds in the post.l/3 of the tongue. - Somatic sensory innervation of the exteroceptors in the external auditory meatus and external ear. |
|
Vagus n. CN-X- |
SVE GVE GVA SVA GSA |
Nucleus ambiguus Dorsal motor nucleus of vagus n. Nucleus of solitary tract Nucleus of solitary tract Spinal nucleus of CN-V- |
- Branchiomotor innervation of laryngeal musculature. - Parasympathetic motor innervation of the thoracic and abdominal viscera. Preganglionic fibres synapse into paramural ggl. (near the viscera) and intramural ggl. (in the walls of the viscera). Postganglionic fibres provide motor parasympathetic innervation for a11 the viscera in the thoracic and abdominal cavities (without pelvic cavity). - Visceral sensory innervation of the interoceptors of laryngeal mucosa and thoraco-abdominal viscera. - Taste of the epiglottis (only in foetus and newborn). - Somatic sensory innervation of the external auditory meatus, external ear and meninges. |
|
Vestibulocochlear n. CN-VIII- |
SSA |
Cochlear and vestibular nuclei in medulla and pons |
- Special sensory fibres, which innervate the auditory and vestibular analysers in the internal ear. |
|
Spinal accessory n. CN-XI- |
SVE |
Spinal accessory nucleus in medulla and cervical spinal cord |
- Branchiomotor innervation for trapezius and sternocleidomastoidian mm. |
|
Hypoglossal n. CN-XII- |
GSE |
Hypoglossal nucleus in medulla oblongata |
- Somatic motor innervation for the muscles of the tongue. |
Conclusion - the internals structure of the brainstem consists of:
grey matter:
nuclei of the cranial nerves,
proper nuclei:
relay nuclei:
motor nuclei:
olivary complex and arcuated nuclei in medulla,
nuclei of the pontine basis (pontine nuclei) in pons,
red nucleus and substantia nigra in midbrain.
sensory nuclei:
gracilis and cuneatus nuclei in medulla,
nuclei of the tectum (tectal nuclei) in midbrain
reticular formation
white matter:
descending tracts (motor tracts) - in the base,
ascending tracts (sensory tracts or "lemnisci") - in tegmentum,
medial longitudinal fasciculus (MLF),
central tegmental tract,
cerebellar afferent and efferent fibres (cerebellar peduncles),
deep unnamed pathways in the reticular formation.
Study of the GREY MATTER
Strategy of study:
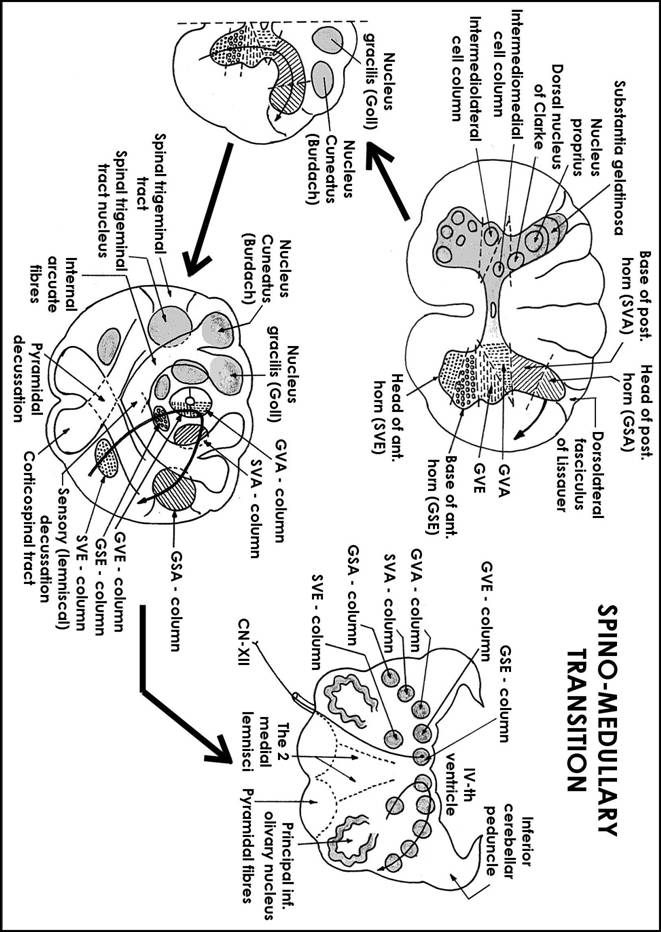
first we shall study the nuclei of CN as a total system at the level of the entire brainstem, because they can be imagined as a prolongation of the spinal grey matter that becomes divided into more isolated masses (nuclei of CN) which will be rearranged in space; this rearrangement respects some steps and rules that coordinate the transition from spinal cord to brainstem;
then we shall study the other nuclei (proper nuclei) and reticular formation, separately, at the level of each segment of the brainstem (medulla, pons and midbrain).
Steps and rules of transition from spinal cord to brainstem:
The most inf. part of the medulla looks very much like the internal structure of the spinal cord; immediately above this level appear the next modifications:
the pyramidal decussation appears; it separates the ant horn of the spinal cord into 2 longitudinal motor columns:
SVE column that corresponds to the head of the ant. horn,
GSE column that corresponds to the base of the ant. horn.
the gracilis and cuneatus nuclei appear; they push antero-laterally the post horn;
the lemniscal decussation (sensory decussation) appears; it separates the post. horn into 2 longitudinal sensory columns:
GSA column that corresponds to the head of the post. horn (thus, substantia gelatinosa becomes spinal trigeminal nucleus and dorsolateral fasciculus of Lissauer becomes spinal trigeminal tract);
SVA column that corresponds to the base of the post. horn
In fact, between GSA and SVA columns appears another column (SSA column which consists of cochlear and vestibular nuclei) and its appearance is not very well explained by this theory; it corresponds to the post. horn (probably to the nucleus proprius);
the components of the intermediate zone of the spinal grey matter (lateral horns) become also separated into 2 longitudinal visceral columns:
GVA column that corresponds to the intermediomedial cell column of the spine
GVE column that corresponds to the intermediolateral cell column of the spine
At the same time with this separation process, the total system (chain) of columns that results migrates after a direction which is:
ventrolateral for those columns which correspond to the post. Horn,
dorsal for those columns which correspond to the ant horn
The migration of the new-formed columns, which is very evident at the level of the sup. 1/2 of medulla (olivary level) is determined probably by the appearance of the olivary nuclear complex. As a result of the migration of the new-formed columns, central canal also migrates posteriorly and expands into the cavity of the IVth ventricle; thus, GSE, GVE and GVA columns become localised into the floor of the IVth ventricle. Their afferent and efferent fibres will interrupt the new-formed longitudinal columns from place to place and thus they become the nuclei of cranial nerves; they are also called equivalent nuclei because they are equivalent to those components of the spinal grey matter from which they result;
GSA column
It corresponds to the head of the post. horn (principally to substantia gelatinosa). It consists of:
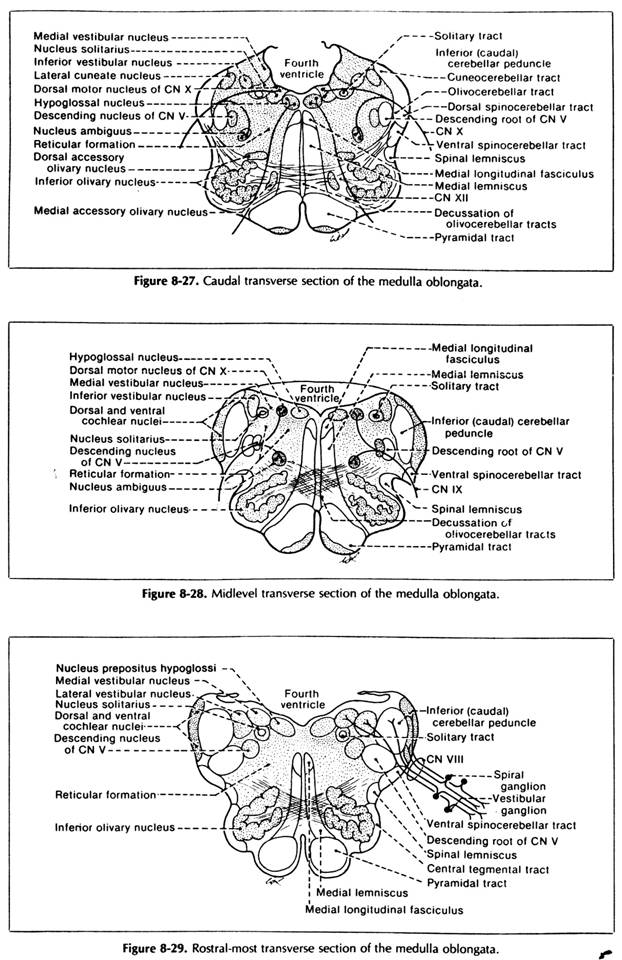
Spinal trigeminal nucleus:
it prolongs rostrally the substantia gelatinosa (they are equivalent nuclei);
it has the most lateral localisation in medulla (it is separated from its lateral surface only by the spinal trigeminal tract);
it presents a somatotopical arrangement of its neurons
pain and thermal stimuli are projected into its caudal part while tactile stimuli are projected into more rostral parts;
fibres of ophthalmic n. project into the ventral part, those of mandibular n. project into the post. part and those of the maxillary n. project in the middle part of the nucleus;
afferent fibres form the spinal trigeminal tract which:
accompanies laterally the nucleus on its whole length,
is the rostral continuation of dorsolateral fasciculus of Lissauer,
its fibres extend between the point of entry of CN-V in pons and the caudal extremity of spinal trigeminal nucleus
Some of the afferent fibres in CN-V bifurcate immediately they enter the pons into an ascending branch (it enters the main sensory trigeminal nucleus in pons) and a descending branch (it enters the rostral levels of spinal trigeminal tract nucleus); they may be considered an equivalent of the medial bundle of fibres in the dorsal root of the spinal nerve; they convey principally the discriminative touch sensation;
efferent fibres (secondary fibres) of this nucleus, together with those from the main sensory nucleus of CN-V (in pons) decussate first and then they ascend forming the trigeminal lemniscus; this lemniscus lies near the medial lemniscus and they both travel to the VPL nucleus of thalamus; other secondary fibres synapse into the reticular formation or establish connections with other nuclei of CN.
Main sensory trigeminal nucleus:
it is localised in the middle level of the pons as a prolongation of the spinal trigeminal nucleus (it lies at the level where the fibres of trigeminal n. enter the pons);
Mesencephalic trigeminal nucleus:
it extends rostrally from the level of entry of the CN-V and reaches the level of sup. quadrigeminal colliculi in midbrain;
in midbrain it lies along the lateral margin of periaqueductal grey matter
it is not a true sensory nucleus because it has no synapses inside it
it only contains the bodies of some primary pseudounipolar neurons (like those of the sensory ganglia);
mesencephalic trigeminal tract consists of the axons and dendrites of these pseudounipolar neurons; the dendrites exit together with the motor root of CN-V and accompany the motor branches to the muscles of mastication where they innervate the central part of muscle-spindles inside masticatory muscles;
the axons of these neurons also contribute to form the mesencephalic trigeminal tract and finally project into the main sensory trigeminal nucleus and into the motor trigeminal nucleus (these last projections usually form reflex connections with the, motor neurons);
Proprioceptive impulses from the muscles innervated by other cranial nerves reach finally also into the mesencephalic trigeminal tract and nucleus.
SSA column
It corresponds to the post. Horn and consists of:
Cochlear nuclei:
they are 2 in number - ventral cochlear nucleus and dorsal cochlear nucleus
they lie on the dorsolateral face of the restiform body (inf. cerebellar peduncle) at the level of medulo-pontine junction;
afferent fibres are represented by the axons of the sensory neurons in the ganglion of Corti; these fibres enter the ventral cochlear nucleus and here they form a band which divides it into an anteroventral and a posteroventral cochlear nuclei; in human, the principal cochlear nucleus is the anteroventral one; neurons in the cochlear nuclei have a tonotopical organisation: fibres from the basal part of cochleea (which bring impulses of high frequency) are projected in the deep parts of the nuclei, while fibres from the apical part of cochleea (low frequency impulses) are projected to the superficial layers of the nuclei;
efferent fibres (second order fibres) behave as it follows:
they may ascend ipsilaterally in the pontine tegmentum forming the ipsilateral lateral lemniscus;
they may decussate forming the trapezoid body and then they also ascend but in the contralateral lateral lemniscus;
both crossed and uncrossed fibres synapse in the sup. olivary complex (formed by medial and lateral sup. olivary nuclei); the function of these nuclei is to localise in space the source of sounds;
lateral lemniscus
is formed by:
efferent fibres of the sup. olivary nuclei,
second order fibres of the cochlear nuclei;
it ascends through the tegmentum of the pons and midbrain and terminates in the inf. quadrigeminal colliculus; efferent fibres from inf. colliculus form the brachium of the inf. quadrigeminal colliculus and ascend to synapse into the medial geniculate body of the thalamus; cortical projections from medial geniculate body terminate in the primary auditory cortex (areas 41-42) in the depth of Sylvian fissure.
Vestibular nuclei:
are 4 in number: sup. one, medial one, lateral one and inf. one;
localisation: in the rostral medulla and caudal pons under area vestibularis of the floor of the IVth ventricle;
afferent fibres are the axons of the neurons in the sensory ganglion of Scarpa; they form the vestibular division of CN-VIII;
efferent fibres
ascending projections
fibres from sup. vestibular nucleus ascend in MLF and project into: trochlear nucleus and oculomotor nuclear complex (its somatic part);
fibres from medial vestibular nucleus also ascend in MLF and project into: extraocular musculature motor nuclei (of CN III, IV and VI), interstitial nucleus of Caja1 (a nucleus at the rostral end of MLF and above oculomotor nuclear complex)
fibres from inf. vestibular nucleus also ascend in MLF and synapses into trochlear and oculomotor complex;
descending projections
fibres from lat. vestibular nucleus form the lat. vestibulospinal tract which descends in the ant funiculus of the spinal cord and synapses in the ant horn, especially at cervical and lumbar levels;
fibres from medial vestibular nucleus descend bilaterally in the MLF and end in the cervical spinal cord;
fibres from inf. vestibular nucleus project to inf. olivary nuclei, medullary reticular formation, cerebellum and cervical spinal cord via MLF.
cerebellar projections
they enter the cerebellum via inf. cerebellar peduncle and project into: flocculo-nodular lobe, uvula, fastigial nucleus (these cerebellar components are totally known as vestibulocerebellum)
reticular projections:
are sent to - nucleus reticularis pontis caudallis and nucleus reticularis gigantocellularis;
cortical projections
they ascend in MLF and terminate into VPL and VPI nuclei of the thalamus; from here they are projected into cortical areas 3a and 2v
SVA column
it corresponds to the base of the post. horn
it is the 'branchiosomatic" this means "branchial sensory") column;
it consists of:
solitary tract nucleus
it lies dorsolateral to the dorsal vagal nucleus;
its general aspect is "tubular" this means that the grey matter, of the nucleus surrounds the white matter of the tract like a tube;
afferent fibres: form the solitary tract and they convey generally impulses concerned with the taste and belong to the cranial nerves VII, IX and X;
GVA column
It corresponds to the intermediomedial cell column of the spinal cord and consists of:
Dorsal sensory nucleus of vagus n.:
it lies dorsomedial to the nucleus of the solitary tract (it represents an extension of the solitary tract nucleus);
it receives the greatest number of vagal afferent fibres (the axons of the sensory neurons in the inf. ganglion of vagus n.) on the way of the solitary tract; some of these afferent fibres synapse into the nucleus of solitary tract; it innervates the carotid body and thoracoabdominal viscera.
Sensory nucleus of glossopharyngeal n.:
it lies in the inf. part of the nucleus of solitary tract
afferent fibres are the axons of the sensory neurons in the inf. ggl. of glossopharyngeal n. and their dendrites innervate the mucosa of the post. 1/3 of the tongue and pharynx; they a1so innervate the baroreceptors of the carotid sinus.
GVA column
It corresponds to the intermediolateral cell column of the spine and consists of:
Dorsal motor nucleus of vagus n.:
it lies in the floor of the IVth ventricle, under the vagal trigone
it provides parasympathetic preganglionic fibres for all thoraco-abdominal viscera except pelvic viscera
Inf. salivatory nucleus:
It is a small nucleus lying near the upper pole of the dorsal vagal nucleus;
It provides parasympathetic preganglionic fibres for parotid gland.
Sup. salivatory nucleus:
it is a prolongation of inf. salivatory nucleus at pontine level;
it provides preganglionic parasympathetic fibres for sublingual, submandibu1ar and lachrymal glands and for the glands of nasal and buccal mucosae;
Edinger-Westphal nucleus:
it lies dorsally to the rostral part of the somatic subdivision of the oculomotor nuclear complex;
it provides preganglionic parasympathetic fibres for the eyeball;
GSE column
It corresponds to the base of the ant. horn of the spine and consists of:
Hypoglossal nucleus:
it lies in the floor of the IVth ventricle at the level of the hypoglossal trigone;
it also extends caudally in the "closed part of the medulla";
its axons run anteriorly between the pyramid and the olive in order to appear in the preolivary sulcus at the surface of the medulla
it innervates the striated muscles of the tongue.
Abducent nucleus:
it lies under the surface of facial colliculus in the floor of the IVth ventricle (its sup. triangle that corresponds to the pons);
- its axons run anteroinferiorly and appear as CN-VI in the bulbo-pontine sulcus above the pyramid; they innervate the lat. rectus m. of the eyeball;
A part of the axons of the abducent nucleus ascend along MLF and reach the somatic subdivision of the oculomotor nuclear complex; they help the conjugated movements of the both eyeballs;
Trochlear nucleus:
it lies immediately caudal to the oculomotor nuclear complex, near the midline, in the tegmentum of the midbrain;
its fibres pass dorsolateral around the periaqueductal grey matter then they decussate to reach the rostral end of sup. medullary vellum; they emerge at the post. surface of the midbrain just laterally to the frenulum of the sup. medullary vellum;
it innervates the sup. oblique m. of the eyeball;
Somatic part of the oculomotor nuclear complex:
it has a "V"-shape with the opening oriented posteriorly
it lies on the midline, ventrally to the periaqueductal grey matter;
it extends between the upper limit of the midbrain and the rostral pole of trochlear nucleus
it is represented by:
lateral somatic cell columns
dorsal nucleus - it innervates the inf. rectus m.;
intermediate cell column - it innervates the inf. oblique m.;
ventral cell column - it innervates the medial rectus m.;
medial cell column - it innervates the sup. rectus m.;
caudal central nucleus - it is a midline somatic cell group in the caudal 1/3 of the complex; it innervates the levator palpebrae superioris m
central nucleus of Perlia - it is a midline cell group concerned with the convergence
axons of the oculomotor nuclear complex pass anteriorly in the midbrain tegmentum, traverse the grey matter of the red nucleus and emerge on the medial face of the ipsilateral crus cerebri.
SVE column
It is a branchiomotor column and corresponds to the head of the ant. horn. It consists of:
Accessory nucleus:
It extends from the upper cervical segments of the spine into the lower medulla, as a prolongation of the central motor column of the ant. horn (phrenic nucleus) - they are equivalent nuclei
its axons emerge on the lat. face of the upper cervical spine as more rootlets which join to form a single trunk that enters the skull cavity through foramen magnum
it innervates the sternocleidomastoidian m. and trapezium m
Nucleus ambiguus:
It extends from the accessory nucleus into the rostral part of the medulla;
its axons, first, run dorsomedially and then they curve laterally to emerge on the lat. face of the medulla; they provide branchiomotor innervation for the stylopharyngeal m. (CN-IX) and laryngeal mm. (CN-X);
Ventrally, in nucleus ambiguus, lies a group of neurons (cardiac vagal nucleus) that innervates the mm. of the heart;
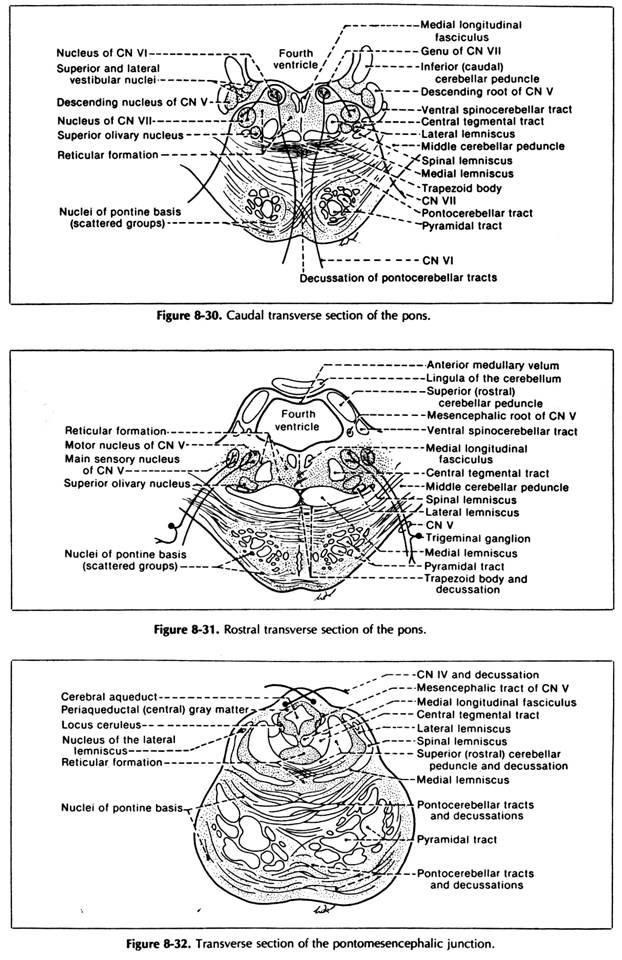
Facial nucleus:
It lies deep in the caudal part of the pontine tegmentum (anteromedial to the spinal trigeminal nucleus)
its axons run:
first dorsomedially and reach medially to the abducent nucleus (here, they lie near the MLF, by which the facial n. can communicate with other cranial nerves);
then the axons surround posteriorly the abducent nucleus (this is the geniculum of the facial n.) forming the facial colliculus in the floor of the IVth ventricle;
finally, they descend anterolaterally through the reticular formation, passing between the facial nucleus and spinal trigeminal nucleus in order to reach in the bulbo-pontine sulcus above the olive;
it innervates the musculature of the facial expression;
Motor nucleus of CN-V:
it lies in the rostral pontine tegmentum near the lateral side of the sup. triangle of the floor of the IVth ventricle
its axons form the motor root of trigeminal n. which takes the way of the mandibular subdivision of trigeminal n.; they innervate the masticatory mm.
PROPER NUCLEI of the Brainstem
Gracilis and Cuneatus nuclei:
localisation - in the dorsal funiculi of the spinal cord that prolong at the level of the "closed part of the medulla"; they lie at the rostral extremities of the tracts with the same names that finish by synapses into these nuclei (the nuclei become greater as the tracts become thinner); cuneatus nucleus lies laterally and slightly rostrally to the gracilis nucleus; cuneatus nucleus also lies dorsally to the reticular formation and medially to the spinal trigeminal nucleus; gracilis nuclei are located on both sides of the median line; both gracilis and cuneatus nuclei determine a round eminence at the level of each corresponding tract, which called tuberculi gracilis and cuneatus
afferent fibres - they receive the fibres of gracilis and cuneatus tracts and also the post. spinoolivary tract which has the fibres intermingled with those of gracilis and cuneatus tracts;
efferent fibres
internal arcuated fibres that decussate and then ascend as the 2 medial lemnisci
projections of the afferent spinoolivary fibres into the inf. accessory olivary nuclei; from here they decussate and form the olivocerebellar tract that projects into the cerebellar cortex;
Accessory Cuneate Nucleus:
it lies dorsolateral to the cuneate nucleus
it contains the same large neurons like those in the dorsal nucleus of Clarke in the spine
afferent fibres - it receives fibres that ascend laterally in fasciculus cuneatus and convey uncounscient proprioceptive impulses from the sup. limb;
efferent fibres - are called post. external arcuated fibres and enter the cerebellum via its ipsilateral inf. cerebellar peduncle;
*** the afferent fibres to this nucleus which are going to be projected to cerebellum are called cuneocerebellar tract
Equivalences that can be established at these levels:
between accessory cuneate nucleus and dorsa1 nucleus of Clarke;
between cuneocerebellar tract and post. spinocerebellar tract;
Nuclei of Trapezoid Body:
They are relay nuclei on the way of efferent cochlear fibres which decussate in order to form the trapezoid body (they lie between the transverse fibres of trapezoid body
Nuclei of the lateral lemniscus:
they are relay nuclei on the way of efferent cochlear fibres (decussated or nondecussated) which ascend as lateral lemniscus to the inf. quadrigeminal colliculi (they lie between the ascending fibres of lateral lemniscus);
*** Both trapezoid body and lateral lemniscus lie in the pontine tegmentum together with their nuclei.
Sup. olivary nuclear complex:
It consists of: medial sup. olivary nucleus and a lat. sup. olivary nucleus
they are also relay nuclei on the way of the auditory fibres in the pontine tegmentum; here synapse some of the efferent cochlear fibres (decussated or not) which are going to ascend in the lateral lemnisci; they are concerned with the space localisation of the source of sounds;
Inf. olivary nuclear complex:
It consists of: principal olivary nucleus medial accessory olivary nucleus, and dorsal accessory olivary nucleus
localisation
principal olivary nucleus lies under the surface of the medullary olive; it practically determines the olivary prominence); it has the general shape of a folded bag whose opening (hillum) is oriented postero-medial;
medial accessory olivary nucleus lies between medial lemniscus (medially), the medullary pyramid (posteriorly) and principal olivary nucleus (posterolaterally)
dorsal accessory olivary nucleus - it lies dorsomedially to the principal olivary nucleus;
efferent fibres
olivocerebellar fibres
arise from all olivary nuclei,
from principal olivary nucleus they arise at the level of its hillum,
they intersect the medial lemniscus, decussate, run postero-laterally and finally enter the cerebellum via its inf. cerebellar peduncle (they are the principal component of this peduncle);
olivospinal fibres
afferent fibres
cortico-olivary fibres
they originate in the cortex of frontal, parietal, temporal and occipital lobes,
they descend together with the corticospinal fibres and synapse bilaterally in the principal olivary nucleus (they are a part of cortico-olivo-cerebellar circuit
fibres from midbrain
they are uncrossed and descend in the central tegmental tract
they are:
rubro-olivary fibres that terminate in the principal olivary nucleus,
fibres from periaqueductal grey matter terminate in the accessory olivary nuclei;
fibres from ponto-medullary nuclei come from: inf. vestibular nuclei, spinal trigeminal nuclei and cerebellar nuclei
fibres from spinal cord: spino-olivary tracts (see ascending tracts of spinal cord)
Pontine Nuclei (Nuclei of the Base of the Pons):
They are many small masses of grey matter scattered between the longitudinal and transverse fibres of the pontine basis. They represent totally the greatest nuclear mass of the pons.
They can be classified into: a lat. nuclear mass, a medial nuclear mass, a dorsal nuclear mass and a ventral nuclear mass.
afferent fibres:
cortico-pontine fibres - they descend from all the regions of the ipsilateral cerebral cortex having a precise somatotopical arrangement; they descend through the base of the midbrain and synapse finally into the pontine nuclei;
efferent fibres
ponto-cerebellar fibres - they are the transverse fibres of the pontine basis; they originate in the pontine nuclei, decussate, and converge laterally in order to form the middle cerebellar peduncle; finally they synapse into the contralateral cerebellar cortex;
As a conclusion we can say that the pontine nuclei are relay nuclei on the cortico-ponto-cerebellar circuit.
Pontine nuclei also receive afferent fibres from the sup. quadrigeminal colliculus (tecto-pontine fibres) which are also transmitted further to the cerebellar cortex (optic afferences of the cerebellum);
Substantia nigra:
It is a bilateral lamina of pigmented neurons (they contain neuromelanin) extending through the entire length of the midbrain between the tegmentum and the corresponding crus cerebri;
its general shape is semilunar with the concavity oriented postero-medially;
rostrally it reaches until the subthalamic region
components
a dorsal part (pars compacta):
it contains numerous neurons in a compact arrangement; they contain a greater amount of pigment
a ventral part (pars reticularis):
it is intermingled with the fibres of the corresponding crus cerebri; for this reason it contains a lesser number of neurons; the amount of pigment is also reduced;
it reaches rostrally the subthalamic region and is considered to be continuous with the globus pallidus;
principal connections with: cerebral cortex, basal nuclei, hypothalamus and spinal cord;
afferent fibres
strio-nigral fibres:
they arise in the - head of the caudate nucleus and putamen;
hey synapse in "pars reticularis" of substantia nigra
subthalamo-nigral fibres
they arise in subthalamic nucleus
they also terminate in "pars reticulata"
tegmento-nigral fibres:
they arise in: midbrain raphe nuclei and pedunculopontine nucleus
they synapse in "pars reticularis" (inhibitory synapses)
efferent fibres:
nigro-striatal fibres
they arise in "pars compacta"
they project into - caudate nucleus and putamen;
nigro-thalamic fibres
they arise in "pars reticularis",
they project into: ventral-anterior nucleus (VA), ventral-lateral nucleus (VL), dorso-medial nucleus (DM)
nigro-tectal fibres
they arise in "pars reticulata"
they project into the middle grey layers of the ipsilateral sup. quadrigeminal colliculus (those portions of the colliculus which are not related with the visual system)
nigro-spinal fibres
they represent a part of the extrapyramidal system and descend in the tegmentum of the brainstem in order to reach the lower motor neurons of the spinal cord;
Red Nucleus:
It is a paired nuclear mass with:
a round shape on transverse section and ovoid shape in longitudinal view
pinkish-yellow colour due to a ferric pigment in its neurons (this colour appears only on fresh material)
central localisation in the midbrain tegmentum (postero-medially to substantia nigra) extending between the caudal margin of the sup. colliculus and caudal diencephalon;
it is traversed from post. to ant. direction by the fibres of the oculomotor nerve in their way to the apparent origin;
it is surrounded at the periphery by a capsule formed by afferent and efferent fibres
afferent fibres
cerebello-rubral fibres
originate in the dentate nucleus of cerebellum,
pass through the sup. cerebellar peduncles,
decussate in the central part of the caudal midbrain tegmentum,
synapse in the contralateral red nucleus
There are also cerebello-rubral fibres that originate in emboliform nucleus and globose nucleus;
*** some of these fibres finish in the reticular formation and thalamus
cortico-rubral fibres
originate in the sensory-motor areas of the cerebral cortex,
they can decussate or not;
vestibulo-rubral fibres
they originate in the contralateral vestibular nuclei,
they mediate the labyrinthic reflexes
other afferent fibres come from - thalamus, subthalamus, hypothalamus, globus pallidus, substantia nigra and spinal cord
efferent fibres
they decussate in the ventral tegmental decussation
they project into:
main sensory nucleus and spinal nucleus of trigeminal n
accessory cuneate nucleus and gracilis and cuneatus nuclei
spinal cord (rubro-spinal tract);
principal inf. olivary nucleus (undecussated fibres)
function of the rubro-spinal fibres - they determine:
excitatory potentials in the contralateral flexor alpha motor neurons
inhibitory potentials in the contralateral extensor alpha-motor neurons
Nuclei of the Tectum:
Inferior Quadrigeminal Colliculus
it has a central ovoid main nucleus surrounded by a laminar zone of nerve fibres - both inf. quadrigeminal colliculi are connected by a commissure of axons of their own neurons
afferent fibres
lateral lemnisci
fibres from medial lemnisci and spinal 1emnisci
efferent fibres
they form principally the brachium of the inf. colliculus and project into the ipsilateral medial geniculate body;
some other efferent fibres project via the sup. colliculi into the next tracts: tecto-spinal tract, medial longitudinal fasciculus and central tegmental tract;
function - they are considered to be a reflex centre for the oculocephalogire reflexes (the turning of the head and eyes to the sources of sounds); this means that they have a role in the localisation of the sources of sounds;
Superior Quadrigeminal Colliculus
It consists of 4 laminae (layers) which from external to internal direction are:
stratum zonale - it consists mainly of fibres coming from the occipital cortex (areas l7, 18, 19); they arrive as cortico-tectal tract; it also contains a few small neurons;
stratum cinereum - it is also called the superficial grey layer; it contains small multipolar neurons on which the cortical fibres synapse partially;
stratum opticum - it is also called the superficial white layer; it consists of fibres of the optic tract which terminate here and after this they are going to invade the adjacent layers
stratum lemnisci - it consists of other 4 layers:
intermediate grey and intermediate white layers represent together the main reception zone; they consist of neurons mixed with axons and dendrites; its afferent source is the medial cortico-tectal pathway from areas 18 (occipital cortex) and area 7 (preoccipital cortex) - these areas are concerned with ocular movements in order to follow different objects);
deep grey and deep white layers contain neurons whose dendrites extend to the optic layer while their axons form many of the collicular efferences;
afferent fibres
from retina (optic tracts) and visual cerebral cortex (occipital lobe); they project into the superficial layers;
from spinal cord (spino-tectal tracts), sensory nuclei (cuneatus, accessory cuneate and spinal trigeminal nuclei), cerebellar nuclei, locus coeruleus, reticular formation of the brainstem and inf. quadrigeminal colliculus; these fibres convey many types of impulses (tactile, thermal, pain, auditory, etc.) and project generally into the deep layers of sup. colliculus; from this point of view, these layers can be compared with the reticular formation having similar functions (they receive and respond to various stimuli);
efferent fibres
they arise in al the layers of the sup. colliculi,
tecto-oculomotor fibres - project into all oculomotor nuclei,
tecto-spinal fibres - descend in the cervical spinal cord and coordinate the movements of the head and neck to the sources of various stimuli (these movements will be conjugated with those of the eyes);
tecto-tegmental fibres - project into the reticular formation substantia nigra and red nucleus;
tecto-geniculate fibres - project into the lat. geniculate bodies of the 2 thalami;
tecto-pontine fibres - they descend to the dorsolateral pontine nuclei (vestibular ones especially) with a relay to the cerebellum;
Descending efferent fibres form the next types of tracts:
tecto-pontine and tecto-bulbar ones which are uncrossed,
tecto-bulbar and tecto-spinal ones which are crossed;
(they form the dorsal tegmental decussation of Meynert);
function:
sup. colliculi are involved in complex integrations between the visual activity and other activities of the body;
Reticular formation:
It represents a special form of existence of the nervous matter which consists of a complex network of white mater (with fibres that run and cross in all the directions) that divides the grey mater into many small, scattered and poorly defined groups of neurons. It fills in the spaces between the nuclei and tracts of the brainstem. It extends on the entire length of the brainstem and continues upwards in the subthalamus and parts of the hypothalamus and dorsal thalamus; downwards it is continued with the reticular formation of the spinal cord;
general characteristics:
its collection of neurons and fibres is located deeply into the nervous matter of the brainstem; they are poorly defined and poorly limited from the rest of the nervous mater; they have diffuse connections
its pathways are complex and polysynaptic; they are very difficult or quite impossible to be defined;
its pathways are both ascending and descending, and in the same time, both crossed and uncrossed (unilateral stimulation often results in bilateral responses);
it contains both somatic and visceral components;
reticular formation can be divided into 3 bilateral longitudinal columns of reticular nuclei
median column
it contains mostly large reticular neurons,
it extends throughout medulla, pons and midbrain,
its nuclei form bilateral vertical sheets that occupy the paramedian zones; they are called the nuclei of the raphe
components:
in medulla (sup.2/3) and ponto-medullary junction lie nucleus raphes obscurus and nucleus raphes pallidus
in pons 1ie nucleus raphes magnus and above it, nucleus raphe pontis; the most sup. localisation has nucleus raphes centralis superior
in midbrain lies nucleus raphes dorsalis (rostralis)
medial column:
it contains generally neurons of medium size, but in some regions they are the largest ones;
in medulla it is unclear in the caudal part (it is represented by a thin lamina lying laterally to the nuclei of the raphe); here it is represented by medullary gigantocellular nucleus (magnocellular nucleus) in the upper medulla; this nucleus lies lateral to the rapheal nuclei, ventral to the Hypoglossal and dorsal vagal nuclei, and dorsal to the inf. olivary complex;
in pons, this column is represented by: pontine gigantocellular nucleus nucleus reticularis pontis oralis and pontis caudalis nuclei cuneiformis and subcuneiformis, and nuclei coeruleus and subcoeruleus which 1ie at the pontomesencephalic junction;
in midbrain there are : dorsal tegmental nuclei deep tegmental nuclei and many other nuclear groups in the periaqueductal grey matter;
lateral column
it contains generally small sized neurons;
components
in medulla central and ventral medullary reticular nuclei (which are confluent and enclose the nucleus ambiguus), Iateral medullary reticular nucleus (equivalent of the nucleus of the lat. funiculus in spinal cord) and paramedian medullary reticular nucleus
in pons pontine parvocellular nucleus which embraces the motor nuclei of facial and trigeminal nerves;
in midbrain lateral and medial parabrachial nuclei pedunculopontine tegmental nucleus-pars compacta (it lies laterally to the decussation of the sup. cerebellar peduncle)
afferent fibres:
from the spinal cord via spinoreticular tracts and collateral branches of long ascending tracts;
from some primary neurons and many secondary neurons of the sensory cranial nerves;
from cerebellum via cerebelloreticular pathways;
from tectal nuclei via tectoreticular pathways which bring indirect information from visual and acoustic pathways;
from thalamic, subthalamic and hypothalamic nuclei
from basal nuclei
from the sensory-motor cerebral cortex and other cortical areas via direct cortico-reticular fibres and collaterals of the corticofugal tracts
from various parts of the limbic system
efferent fibres
to the somatic motor and visceral motor centres of the spinal cord via reticulospinal tract,
to the somatic motor and visceral motor centres of the brainstem via short descending pathways;
to the cerebellum
to the red nucleus, substantia nigra and tectal nuclei
to the subthalamus, thalamus and hypothalamus
indirectly to the basal nuclei and cerebral cortex (neocortex and limbic system) via irradiations of the diencephalic nuclei;
functions
somatomotor control - reticular formation is a part of a complex of subcortical regions influencing the activity of the somatic musculature, by the influence exerted on alpha- and gamma-motor neurons (reticulospinal tracts)
somatosensory control - all the ascending pathways from the spinal cord or brainstem are under the facilitator or inhibitory control of the reticular formation (many of these pathways have fibres or collaterals which synapse into reticular formation);
visceromotor control - cardiovascular activity and the activity of the other thoraco-abdominal viscera are either directly or indirectly under the partial control of reticulo-bulbar and reticulo-spinal fibres; thus appeared the concept of "cardiovascular centre" or "respiratory centre
neuro-endocrine transduction - it is exerted via the connections between reticular formation and hypothalamus;
biological rhythms - the "biological clock" involves the rhythms of different biological activities like reproductive cycles, development, cell division, cell death and replacement and many others; many rhythms depend on an intact hypothalamus which, as we know, has many connections with reticular formation; in the same way the "circadian rhythms" (day-night rhythms) can also be influenced;
sleep, arousa1, states of consciousness, perception - arousal and awakening are depending on an ascending activating reticular system which:
has many inputs from the ascending pathways,
has polysynaptic relays in reticular formation,
has diffuse and widespread projections in both cerebral hemispheres at the level of the neocortex and limbic system;
spatio-temporal discrimination, learning and memory, cognitive and exploratory function, emotions and long-term homeostasis - all depend on an intact hypothalamo-limbic system which receives widespread impulses from numerous reticular elements;
Study of White Matter
It represents the system of descending fibres that occupies the base of the brainstem. It conveys voluntary motor impulses, formed generally in the primary motor somatic area.
It can be classified into: corticospinal fibres, corticopontine fibres, corticobulbar fibres and corticonuclear fibres
corticospinal fibres:
they descend through the internal capsule (they occupy a position near its genu and extend into its post. limb);
they reach at the level of each crus cerebri on the same side where they occupy the middle crural 2/3 together with the corticonuclear fibres;
at the level of the pons they form groups of bundles at the level of its base but they also remain ipsilaterally;
at the level of the medulla they represent 2 great bundles of fibres that form the 2 medullary pyramids; they are going to form the pyramidal decussation at inferior medullary level in order to extend into the spinal cord (see the tracts of the spinal cord);
corticopontine fibres
they form 2 groups of fibres:
frontopontine fibres
they arise in the frontal cortex (areas 4 and 6 principally);
they descend together with the corticospinal fibres in the internal capsule;
they occupy the medial 1/6 of each crus cerebri, ipsilaterally
temporopontine fibres- they arise in the temporal lobe cortex - they reach in the lateral 1/6 of each crus cerebri ipsilaterally;
parieto- and occipitopontine fibres are also described in each crus cerebri, lying medially to the temporopontine fibres;
all the corticopontine fibres finish by synapses into the pontine nuclei in the ipsilateral half of the pontine base (see the , motor relay nuclei of the brainstem); the axons of the neurons in the pontine nuclei form the transverse pontine fibres that decussate and converge laterally in order to form the middle cerebellar peduncles and finally project into the contralateral cerebellar cortex;
corticobulbar fibres
they descend together with the corticospinal fibres until they reach the medullary pyramids;
in the pyramids they synapse into the arcuate nuclei which lie on the ant. aspect of each pyramid;
axons of the neurons in the arcuate nuclei form the ant. external arcuate fibres which will decussate and surround the surface of the contralateral half of the medulla in order to enter the contra lateral inf. cerebellar peduncle;
they are a part of the cortico-bulbo-cerebellar circuit;
corticonuclear fibres (cnf)
they are the equivalent of the corticospinal fibres for the nuclei of the brainstem;
they generally travel in the company of corticospinal fibres until they reach the neighbourhood of the target nucleus; here they leave the corticospinal fibres and enter the tegmentum of the brainstem in order to synapse with the neurons of the target nuclei; these nuclei are:
somatic motor nuclei of the cranial nerves,
nuclei of the reticular formation in the adjacent places;
cnf for oculomotor nuclei leave the crurae cerebri and enter the midbrain tegmentum in order to innervate the somatic part of the oculomotor nuclear complex, trochlear and abducent nuclei; these nuclei are responsible for the innervation of the extraocular mm. of the eyeball and levator palpebrae superioris m.
cnf for SVE nuclei of CN-V (motor trigeminal nuclei) will leave the corticospinal tract near the border between midbrain and pons and enter the pontine tegmentum in order to innervate these nuclei which are responsible for the innervation of all the masticatory muscles; they receive bilateral projections from cortical upper motor neurons except external pterygoid mm. which will receive this innervation only from the contralateral somatomotor cerebral cortex;
cnf for SVE nuclei of CN-VII (motor facial nucleus) enter the tegmentum at the level of the midpons; the lower motor neurons which innervate the muscles of the sup. 1/2 of the face receive bilateral projections from the upper motor neurons while those innervating the lower 1/2 of the face receive upper motor neuron innervation only from the contralateral somatomotor cerebral cortex;
cnf for SVE nucleus of CN-IX and CN-X (nucleus ambiguus) leave the corticospinal tract at the level of lower pons or upper medulla; they project bilaterally into lower motor neurons of ambiguus nuclei; they innervate the mm. of pharynx and larynx;
cnf for SVE nucleus of CN-XI (spinal accessory nucleus) leave the corticospinal fibres in the midlevel of medulla and project ipsilaterally to these nuclei; they innervate the sternocleidomastoidian and trapezium mm.
cnf for GSE nuclei of CN-XII (hypoglossal nucleus) leave the corticospinal tract in medulla, pass through the medullary reticular formation and project into hypoglossal nuclei bilaterally; all the muscles of the tongue receive bilateral upper motor neuron innervations except genioglossus m. which receives upper motor neuron innervation only from the contralateral cerebral somatomotor cortex;
Normally, the reticular formation plays a role in:
the modulation of segmental and postural reflexes and
the regulation of the muscle tone
The brainstem reticular formation plays also a prominent role in the control of the voluntary motor activity. Corticoreticulospinal system consists of 2 types of fibres: corticoreticular and reticulospinal; at the same time this system includes the nuclei of the brainstem reticular formation on which the corticoreticular fibres synapse;
corticoreticular fibres
arise from:
the premotor cortex (most of the area 6 of the precentral gyrus);
the supplementary motor area (the medial and superolateral portion of the area 6);
descend through the base of the brainstem;
project into some of the neurons of origin of the reticulospinal tracts; thus, some of the reticular neurons of origin of reticulospinal tracts are under the influence of cortical neurons while the others are not;
reticulospinal fibres
form a medial and a lateral reticulospinal tract (see the descending tracts of the spinal cord);
cortically driven neurons whose axons descend into the medial reticulospinal tract have principally inhibitory function upon the spinal interneurons and motor neurons which are concerned with the innervation of the axial musculature of the trunk and the muscles of the limb girdles; they inhibit the stretch reflexes allowing the smooth, and rapid execution of the voluntary motor actions;
cortically driven neurons whose axons descend into the lateral reticulospinal tract modulate generally the activity of the spinal motor neurons which innervate the more distal limb muscles;
as a conclusion we can say that the corticoreticulospinal system has a complementary role to the motor function commanded by the corticospinal system; the corticospinal system provides the signals for the movements and the corticoreticulospinal system provides the signals for other muscles which are required in order to execute the principal motor action and, at the same time, it blocks the segmental reflexes (like stretch reflexes for example) which might interfere with the smooth execution of the principal motor action;
It is a paired ascending tract with the next course and relations
At medullary level
it begins immediately above the level of the pyramidal decussation in medulla. It is formed by the axons of the neurons in the nuclei gracilis and cuneatus; these fibres run first antero-medially as internal arcuate fibres which cross the midline forming the lemniscal decussation which represents practically the beginning of the 2 medial lemnisci
lemniscal decussation lies posteriorly to the pyramids and anteriorly to the central grey matter of medulla; the decussation pushes practically more posteriorly the central grey matter and central canal which is going to become the fourth ventricle;
the 2 medial lemnisci ascend on both sides of the median raphe as 2 flattened tracts which have triangular shape on transverse section; the 2 triangular shapes have the longest dimension on the antero-posterior direction at medullary level;
Relations of the lemnisci at medullary level
ant. - the pyramids,
post. - the medial longitudinal fasciculi and tectospinal tracts,
lat. - the inf. olivary nuclear complex,
medial - the median raphe
Somatotopical arrangement in the lemnisci:
fibres of the gracilis tract lie laterally and those of cuneatus tracts lie medially (they are rearranged);
At the pontine level the 2 medial lemnisci have a ventral localisation in tegmentum; their shape is of a flattened oval with the great axe oriented transversally; they also extend laterally beginning from the median raphe of the pons; their vertical fibres are traversed by the horizontal axons which will form the trapezoid body (trapezoid body represents the decussation of the efferent fibres of the cochlear nuclei and it lies ventrally to the lemniscal fibres and dorsally to the base of the pons; further, these fibres will form the lateral lemnisci); the medial lemnisci are related laterally with other lemnisci: trigeminal lemnisci, spinal lemnisci lat. spinothalamic tracts), and lateral lemnisci; all these lemnisci form together a transverse bundle of fibres on transverse section; as they ascend, the lateral lemnisci reach near the dorsolateral face of the brainstem;
At the midbrain level all the lemnisci described until now form a curved band postero laterally to the substantia nigra; the concavity of the curved band (on transverse section) is oriented posteromedially;
Lateral lemniscus - see the efferent fibres of the cochlear nuclei and the relations of the medial lemnisci;
Spinal lemniscus
It is represented by the fibres of the spinothalamic tracts which travel at the level of the brainstem:
at medullary level it travels as a thin bundle located at the lateral surface of the medulla, anteriorly to the spinocerebellar tracts.
for the relations at the level of the pons and midbrain see the relations of the medial lemniscus;
Medial longitudinal fasciculus (MLF):
It is a composite tract made by high myelinated fibres. Its course, relations and function:
at the midbrain level it lies ventrolaterally to the oculomotor nuclear complex; it ascends to the interstitial nucleus of Cajal in the lat. wall of the IIIrd ventricle, where it finishes;
-at the levels of the pons and medulla, MLF maintains generally its ventral position to the central grey matter; in medulla it will be pushed more posteriorly by the decussations of the corticospinal tracts and of the medial lemnisci;
on its way, MLF is closely related with: oculomotor, trochlear and abducent nuclei, emerging facial fibres, dorsal cochlear fibres and hypoglossal nucleus; this fact transforms MLF into a convenient interconnecting route in the brainstem, between all these structures; - it also contains fibres from lateral lemniscal nucleus and from the vestibular nuclei; thus it has the function to coordinate the movements of the eyes and head as an answer to vestibulo-cochlear stimulation;
it is thought that the coordination between the facial and hypoglossal nerves in the movements of the tips and tongue in speech attributed to the connections via the MLF;
vestibular fibres ascend or descend via MLF reaching on its way to the nuclei of CN III-VI and XI, interstitial nuclei (Cajal), nuclei of the post. commissure, and to the spinal cord (medial vestibulospinal tract); some vestibular fibres may ascend to the thalamus on the way of MLF;
Central tegmental tract
At the midbrain level it has a "central tegmental localisation; here it lies:
laterally to the MLF,
dorsolaterally to - the red nucleus and the decussation of the sup. cerebellar peduncles,
At the pontine level and medullary level it deviates laterally to reach and finish into the inf. olivary complex;
It contains both ascending and descending fibres;
descending fibres come from: thalamus, periaqueductal grey matter, red nucleus and basal ganglia; it represents a pathway from the motor cerebral cortex to the brainstem reticular nuclei and inf. olivary complex;
*** Both ascending and descending fibres in this tract might belong to the brainstem reticular formation, their collaterals and terminals innervating other reticular or adjacent specific nuclei.
|
