Radiatiile
pe care le numim neionizante sunt radiatii de natura
electromagnetica cu lungimea de unda ![]() cuprinsa în domeniul 150 nm si
undele centimetrice.
cuprinsa în domeniul 150 nm si
undele centimetrice.
Radiatiile electromagnetice reprezinta o forma de propagare în spatiu a energiei unui câmp electric E si a unui câmp magnetic B, ai caror vectori oscileaza cu aceeasi perioada T, sau frecventa n, în plan perpendicular unul pe celalalt si care sunt perpendiculare pe directia de propagare a undei cu viteza "v".
Orice radiatie electromagnetica poate fi considerata simultan atât unda cât si corpuscul (foton). In functie de frecventa undelor, se manifesta cu precadere fie unul, fie celalalt aspect. La frecvente joase (unde radio, TV, microunde) predomina caracterul ondulatoriu, iar pentru radiatiile cu frecventa mare (infrarosii, vizibile, ultraviolete) începe sa se manifeste si caracterul corpuscular.
Pentru explicarea interactiei radiatiilor electromagnetice cu substanta (efect fotoelectric, reactii fotochimice, împrastiere), se impune considerarea caracterului corpuscular (flux de fotoni), principalul fenomen fizic rezultat în urma acestei interactiei fiind absorbtia de energie.
Din categoria radiatiilor neionizante ne vom ocupa de: radiatiile ultraviolete cu l între 150-400 nm, radiatiile vizibile cu l între 400-800 nm, radiatiile infrarosii cu l între 800-106 nm, microundele cu l între 106-109 nm.
Absorbtia radiatiilor electromagnetice de catre atomi determina tranzitii ale electronilor, de pe nivelele energetice de baza, pe nivele mai ridicate, trecând astfel din starea fundamentala în stari excitate. Daca cuanta de energie absorbita (cantitatea transportata în intervalul unei perioade de oscilatie) este superioara energiei de legatura a electronului în atom, electronul va fi smuls, atomul devenind ion pozitiv 646e46g . Valorile cuantelor de energie ale radiatiilor UV, vizibile, IR si ale microundelor sunt mai mici decât energia necesara ionizarii atomilor (energia de ionizare este de 13,34 eV pentru hidrogen, de 13,57 eV pentru oxigen, de 11.24 eV pentru carbon, de 14,51 eV pentru azot, în timp ce energia radiatiilor UV este cuprinsa între 12,4-3,1 eV, a radiatiilor vizibile între 3,1-1,65 eV, a radiatiilor IR între 1,65-1,2·10-3eV, iar pentru microunde între 10-3-10-5eV.
In cazul moleculelor formate din cel putin doi atomi, trebuie sa tinem seama ca nivelele energetice ale unei molecule nu sunt date (ca în cazul atomilor) numai de valorile energiei electronilor (Eel), ci si de energia de vibratie (Evibr) si de energia de rotatie (Erot) a atomilor din molecula.
Valorile Evibr si Erot sunt tot cuantificate, ca si Eel, iar între aceste nivele sunt posibile mult mai multe tranzitii decât în cazul nivelelor electronice. Desi figura nu reprezinta cazul unei anumite molecule, trebuie remarcat faptul ca diferentele între nivelele electronice sunt mai mari decât între nivelele de rotatie sau vibratie. De aceea, tranzitiile între nivelele electronice se fac cu absorbtia sau emisia unor cuante de energie corespunzând radiatiilor UV si vizibile; tranzitiile vibro-rotationale corespund domeniului IR iar tranzitiile de rotatie pura corespund fie domeniului IR îndepartat, fie microundelor. DEel este de ordinul 1eV, DEvibr este de ordinul 10-1eV, iar DErot este de ordinul 10-2eV.
In ansamblu, pentru o molecula ca cea descrisa:
![]()
Se stie ca într-un atom (sau molecula) pot exista doi electroni care sa ocupe simultan acelasi nivel energetic (sa se gaseasca în acelasi orbital), cu conditia ca acestia sa aiba spinul opus (prin spin se întelege momentul cinetic propriu al electronului datorita rotatiei în jurul axei proprii. Spinul electronic poate avea numai doua valori egale în modul dar opuse ca semn).
Atunci când un electron absoarbe o cuanta de energie electromagnetica, el trece pe un nivel energetic superior (este excitat) si în mod normal nu-si schimba starea spinului. Respectiva stare excitata este numita în acest caz singlet.
Desi cu probabilitate mult mai mica, este totusi posibil ca odata cu excitarea, electronul sa-si inverseze si spinul, respectiva stare numindu-se triplet.
Timpul de viata al starilor excitate este în general foarte mic (10-9÷10-8s), datorita tendintei spontane a electronului de a reveni în starea fundamentala. Deoarece tranzitia din starea triplet în starea fundamentala înseamna iarasi o inversare de spin, timpul de viata al starii triplet este mai mare (între 10-5 si câteva secunde) decât cel al starii singlet. In acest interval de timp mai lung este posibil ca molecula excitata sa transmita surplusul de energie altei molecule (prin ciocniri) si sa îl emita sub forma de radiatie electromagnetica.
Dezexcitarea moleculei se poate face deci prin tranzitii neradiative si, respectiv, tranzitii radiative fluorescenta, fosforescenta):
Fluorescenta se realizeaza atunci când molecula revine în starea singlet fundamentala dintr-o stare excitata tot singlet.
Fosforescenta se realizeaza atunci când molecula revine în starea singlet fundamentala dintr-o stare excitata triplet.
In cazul tranzitilor neradiative, energia fotonului absorbit este fie convertita complet în caldura (agitatie termica) prin ciocniri, fie transferata partial altei molecule, capabila sa treaca, la rândul ei într-o stare excitata.
Moleculele de importanta biologica (proteine, acizi nucleici etc.) contin de regula, un mare numar de atomi, astfel ca, în acest caz, ca urmare a absorbtiei radiatiei de catre acestea, sunt posibile foarte multe tranzitii între diferitele nivele de energie: electronice, de vibratie si de rotatie. De aceea spectrele de absorbtie ale acestor molecule nu contin linii spectrale distincte ca în cazul atomilor izolati, ci grupuri de benzi. Pe de alta parte atunci când aceste molecule se gasesc în solutie sau în contact strâns cu alte molecule, asa cum este cazul în celule, rotatia moleculelor si chiar vibratiile sunt modificate de prezenta celorlalte molecule, efectul fiind o suprapunere a nivelelor energetice în asa masura încât în loc de linii sau benzi de absorbtie, spectrul de absorbtie devine continuu.
Radiatiile electromagnetice cu ![]() între 150 - 800
nm sunt radiatii care activeaza moleculele. Radiatia
absorbita poseda energie suficienta pentru a determina
transformarea moleculelor pe baza unor reactii fotochimice. Principalele
tipuri de reactii fotochimice sunt:
între 150 - 800
nm sunt radiatii care activeaza moleculele. Radiatia
absorbita poseda energie suficienta pentru a determina
transformarea moleculelor pe baza unor reactii fotochimice. Principalele
tipuri de reactii fotochimice sunt:
Excitarea moleculelor M ----- ----- ---->M*
Reactii fotochimice M*+A --------->D
Reactii de dimerizare M*+M -------->MM
Reactii de fotosensibilizare M*+N -------->M+N*
Prima relatie reda excitarea moleculei initiale, a treia si a patra relatie sunt cazuri particulare de reactii fotochimice.
Se defineste randamentul fotochimic ca raport între numarul de molecule transformate si numarul de cuante de energie absorbite.
Randamentul este subunitar, radiatia absorbita nu duce în mod obligatoriu la o reactie fotochimica (absorbtia fotonilor constituie numai etapa primara, urmata de o succesiune de alte etape secundare ce conduc la transformarea fotochimica propriu zisa.
Desi procesele fotobiologice sunt foarte diverse si înca incomplet elucidate, pentru unele din ele se pot distinge urmatoarele stadii:
Absorbtia cuantelor radiatiilor;
Procese intramoleculare cu schimb de energie;
Transportul energiei starii excitate între molecule (migratia energiei);
Reactia fotochimica primara;
Modificarile la întuneric ale produsilor fotochimici primari, care se sfârsesc cu formarea unor produsi stabili;
Reactii biochimice cu participarea fotoprodusilor;
Raspunsul fiziologic celular.
Procesele fotobiologice încep cu absorbtia radiatiilor incidente de catre compusi specifici, specializati, biologici si se încheie cu initierea unei reactii fiziologice din partea sistemului biologic.
Se disting procese fotobiologice cu efecte negative si efecte pozitive:
Efectele negative în organismele vii se împart în doua tipuri:
efecte fototoxice
Efectele fototoxice sunt modificarile produse de radiatii la nivelul pielii si ochilor. Acestea declanseaza reactii fiziologice reactive care se manifesta clinic sub forma de eritem, edem, pigmentatie.
Aparitia eritemului si a pigmentatiei se datoreaza absorbtiei radiatiilor cuprinse între 240-320 nm, pentru eritem si între 300-420 nm pentru pigmentatie. Se presupune ca mecanismul molecular în aparitia eritemului ar fi lezarea termica a fosfolipidelor ce intra în constitutia membranelor biologice. Pigmentatia este realizata de celule speciale din piele, melanocite, care cu ajutorul unei enzime, tirozinaza, transforma tirozina în melanina (pigment ce absoarbe 90% din radiatiile UV).
Asupra ochilor, radiatiile UV produc o reactie inflamatorie a corneei, de aceea se recomanda, pentru protectie, folosirea unor ochelari din sticla speciala.
efecte fotoalergice
Efectele fotoalergice declanseaza mecanismele imunologice primare de sensibilizare alergica, ce se manifesta mai ales prin edeme inflamatorii, eritem, prurit.
Efectele pozitive sunt:
Fotosinteza, fototropism, fototactism (la plante) si fotoperiodism.
Fotoreceptia la nivelul celulelor senzitive retiniene.
Formarea vitaminei D2.
Eritemul caloric.
Fotosinteza, ca mecanism, a fost descrisa anterior. O serie de plante prezinta fenomenul de fototropism, fenomen ce se concretizeaza prin tendinta unor plante de a se orienta în directia sursei de lumina. Fenomenul de fototactism se refera la tendinta de apropiere sau de departare de sursa de lumina.
Fotoperiodismul consta în reglarea ciclului de viata al organismelor vii (plante sau animale) pe baza actiunii ciclice lumina-întuneric. Procesul se petrece sub actiunea luminii vizibile.
Mecanismul molecular al fotoreceptiei senzoriale
Receptorii vizuali, celulele cu bastonas, reprezinta un exemplu de celula receptoare la care a fost elucidata în detaliu succesiunea de procese moleculare prin care stimulul (radiatii vizibile cu l între 400 si 800 nm) determina aparitia potentialului de receptor.
Celulele receptoare din retina sunt de doua tipuri: celule cu conuri si celule cu bastonas. Conurile sunt responsabile cu vederea diurna sau fotopica, iar bastonasele de vederea crepusculara (scotopica). Segmentul extern de bastonas din retina, specializat în fotoreceptie, contine circa 1000 de discuri suprapuse unul peste celalalt. Fiecare disc este un organit separat în citoplasma. Discurile sunt spatii închise delimitate de o membrana foarte fluida având o proportie foarte mare de fosfolipide si foarte putin colesterol.
Discul contine 60% proteine si 38% lipide, iar rhodopsina reprezinta 80% din totalul proteinelor. Rhodopsina se gaseste în membrana discurilor si în membrana citoplasmatica a segmentului extern de bastonas, dar nu si în restul membranei citoplasmatice a acestui tip de celula. Rhodopsina contine o parte proteica opsina si o grupare prostetica numita retinol, atasata specific într-un loc hidrofob pe proteina. Rhodopsina este insolubila în apa. Ea reprezinta proteina fotoreceptoare din retina. O celula cu bastonas este excitata de un singur foton. Sub actiunea acestuia are loc izomerizarea retinolului din forma 11-cis în forma 11-trans, ceea ce schimba foarte mult geometria acestuia. El nu se mai potriveste steric în locul de legare de pe proteina.
Apar modificari conformationale ale proteinei, ce au ca urmare desfacerea retinolului de opsina si descompunerea rhodopsinei. Procesul este reversibil la întuneric.
Membrana citoplasmatica a segmentului extern de bastonas contine numeroase canale de Na+, care în conditii de întuneric sunt deschise, permitând ionilor de Na+ sa intre, ca urmare a potentialului lor electrochimic mare din mediul extracelular. Ionii de Na+ intrati nu se acumuleaza în celula, deoarece sunt evacuati continuu de pompele ionice din membrana corpului celulei receptoare, urmând sa reintre în bastonas prin canalele de Na+. Se creeaza astfel un curent electric transportat de ionii de Na+, numit curent de întuneric. Potentialul electric intracelular este de -20 mV în raport cu potentialul zero al mediului extracelular. Sub efectul luminii, rhodopsina se activeaza si determina închiderea canalelor de Na, cu disparitia curentului de întuneric si hiperpolarizarea celulei al carei potential devine mai negativ (-35 mV). Aceasta variatie este suficienta pentru a declansa excitatia neuronilor bipolari. Potentialele locale aparute în neuronii bipolari determina excitarea neuronilor ganglionari, care genereaza PA, ce ajung la nivelul scoartei unde produc senzatia vizuala. Faptul ca este suficienta o singura cuanta de lumina pentru a produce excitarea receptorului, în timp ce energia necesara hiperpolarizarii este cu mult mai mare, sugereaza ca modificarea spatiala a moleculei de rhodopsina reprezinta doar declansarea unor procese de recrutare energetica. Aceste procese consumatoare de energie metabolica determina aparitia potentialelor locale a caror amplitudine este proportionala cu intensitatea luminii incidente. Variatiile locale de potential membranar, sunt preluate si însumate de celulele bipolare retiniene. PA a caror frecventa este proportionala cu intensitatea stimulului, apar numai la nivelul celulelor ganglionare, respectiv în fibrele nervului optic.
O alta reactie fotochimica deosebit de importanta pentru orgasnism, si care se produce în piele în urma absorbtiei radiatiilor ultraviolete, este reactia de transformare a ergosterolului în vitamina D2. Spectrul de actiune are un maxim la 280 nm.
Actiunea biologica a radiatiilor infrarosii (eritemul caloric) este consecinta absorbtiei lor de catre celulele pielii si a tesutului subcutanat (putere mica de patrundere). In urma absorbtiei radiatiilor infrarosii, pielea si tesutul subcutanat se încalzesc, ceea ce face sa creasca metabolismul local si sa se produca, de asemenea, o vasodilatatie locala. Aceste radiatii, în functie de doza absorbita si de starea pacientului, pot fi folosite în tratamentul unor afectiuni, cum ar fi: edeme inflamatorii, celulite, tulburari de circulatie periferica etc.
RADIAŢII IONIZANTE
radiatiile electromagnetice cu l<100Å si energie >100 eV
radiatiile X si g
radiatiile corpusculare
radiatiile a, b (pozitroni) si b-(electroni)
protoni,
deuteroni,
neutroni rapizi,
ioni grei accelerati,
fragmente de fisiune.
Radiatiile corpusculare si g îsi au originea în nucleul atomic, iar radiatiile X provin din straturile electronice profunde (a se vedea cursul de Radioimagistica).
Fenomenul fizic principal care sta la baza emisiei de radiatii nucleare este dezintegrarea radioactiva. Aceasta consta în scindarea spontana a nucleelor instabile (izotopi/nuclizi) cu formarea unui nucleu mai usor, sau cu formarea a doua nuclee mai usoare (fisiune nucleara). Este un proces spontan, care se desfasoara cu o anumita probabilitate bine determinata pentru fiecare nuclid în parte.
energia transportata de radiatiile ionizante în mediile pe care le strabat este mult mai mare decât energia care leaga orice electron în atom sau atomii între ei.
genereaza fenomene de ionizare (directa sau indirecta) a atomilor sau moleculelor mediilor pe care le strabat.
produc excitari atomice si moleculare
produc radicali liberi.
Aspecte fizice ale interactiei cu materia
Radiatiile corpusculare
Interactioneaza direct cu atomii si moleculele mediului strabatut, probabilitatea de interactiune fiind maxima cu electronii orbitali (învelisul electronic al atomului) producând un numar mare de ionizari. Radiatiile b (electroni sau pozitroni accelerati) pot interactiona si cu nucleele. În câmpul coulombian al unui nucleu greu, electronul este frânat, o parte din energia sa transformându-se în radiatie electromagnetica X de frânare (vezi cursul de Radioimagistica).
Neutronii nu au sarcina electrica si nu interactioneaza cu învelisul electronic al atomilor. Pot interactiona cu nucleele, iar efectele sunt diferite functie de viteza acestora (energia). Neutronii rapizi (E > 1 KeV) produc ciocniri elastice care pot duce la dislocarea nucleelor mici (de ex. H) si astfel determina fenomene de ionizare indirecta. Neutronii lenti sunt absorbiti de nucleu si determina excitarea acestora cu emisie secundara de fotoni g la dezexcitare.
Interactiunea
radiatiilor ionizante de natura electromagnetica cu materia se
face cu electronii liberi din mediu, prin trei tipuri de efecte cunoscute: efect fotoelectric, efect
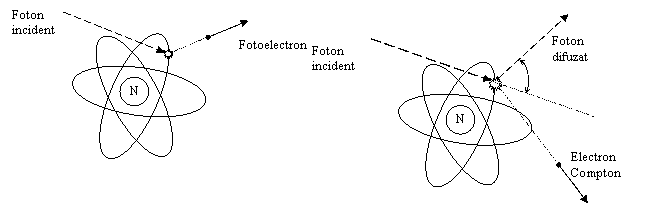
 Efect
fotoelectric
Efect
fotoelectric
Efect Compton
Generare de perechi
E > 1,22 MeV
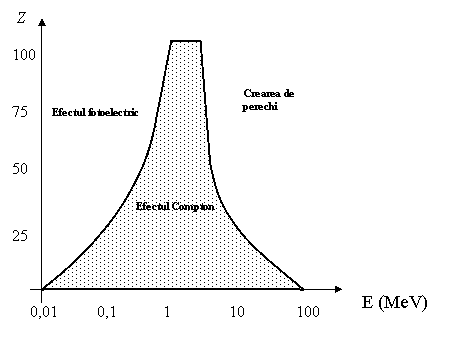
Efectele depind de numarul atomic Z si de energia radiatiei:
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Parcurs = grosimea de material care reduce energia particulei la zero.
Densitatea de ionizare = numarul de perechi de ioni create pe unitatea de parcurs.
Transferul Liniar de Energie (TLE) = raportul dintre energia cinetica a particulei si lungimea traiectoriei în mediul dat.
Este proportional cu numarul de ionizari produse pe unitatea de lungime (N) si cu energia transferata la fiecare ionizare (e):
TLE = N · e
Traiectoria pentru particulele grele (a, protoni) traiectoriile sunt rectilinii, întrucât masa lor mare le confera o inertie suficient de mare pentru a nu fi deviate în momentul ciocnirii cu un electron. Radiatiile b (electroni de mare viteza) au traictorii mai sinuoase, din cauza multiplelor devieri pe care le sufera. Parcursul lor este mult mai lung din cauza unui TLE mult mai mic.
Proprietati specifice radiatiilor fara sarcina electrica (electromagnetice si neutroni)
Traversarea de catre fascicul a unui mediu dat, a carui grosime este x, se face printr un proces de atenuare a carui lege este (Langmuir):
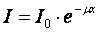
unde I este intensitatea fasciculului initial, I este intensitatea fasciculului dupa strabaterea stratului de grosime x, iar m este coeficientul de atenuare caracteristic substantei traversate si dependent de energia radiatiei.
Radiatiile electromagnetice nu sunt niciodata complet absorbite. Au grad de penetrabilitate foarte ridicat. Grosimea de înjumatatire (x1/2) este distanta la care I I
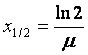
Astfel, radiatiile X si g sunt înjumatatite de un strat de 100 m în aer si de un strat de apa de 10 cm. Pentru 60Co (Eg = 1,3 MeV), x1/2 = 11 mm în Pb.
Efectele moleculare ale radiatiilor ionizante
Radiatiile produc efecte biologice la nivel molecular prin actiune directa, indirecta si la distanta.
a) efectul direct
Absorbtia energiei radiatiilor duce la excitarea si ionizarea atomilor si moleculelor componente ale celulelor. Acesta reprezinta efectul primar fizico-chimic. Toate componentele celulare, inclusiv proteinele citoplasmatice (enzimele), materialul cromozomic (nucleoproteidele sau compusii DNA cu proteinele) si moleculele de apa, sufera actiunea directa a radiatiilor. Efectele directe determina rupere de legaturi covalente sau coordinative cu denaturarea structurala si aparitie de noi compusi chimici (radioliza apei).
b) efectul indirect
Ansamblul de modificari si transformari chimice pe care le sufera moleculele care au fost excitate sau ionizate de radiatii. Este secundar, datorat în special radicalilor liberi proveniti prin radioliza apei continute în tesut.
Radioliza apei depinde de natura radiatiei, de energia ei si de prezenta oxigenului (în lipsa oxigenului radiatiile X si g nu descompun apa).
Radicalii formati sunt deosebit de reactivi, au durata de viata scurta si sunt capabili sa declanseze reactii biochimice în lant, ceea ce determina amplificarea leziunilor directe.
In urma interactiei, molecula de apa pierde un electron si se transforma în ion pozitiv.
H2O H2O+ + e-
Electronul este captat de o alta molecula de apa, ce se transforma în ion:
H2O + e- H2O-
Ionii formati sunt instabili, se descompun imediat dând nastere la radicali liberi si doi ioni stabili care recombinându-se dau molecula de apa:
H2O H+ + O·
H2O-
H+
+
Radioliza apei în sine nu determina un efect biologic direct. Numarul de molecule care sufera actiune directa este extrem de mic în raport cu numarul total de molecule prezente. Iradiind cu o doza de 1 R, un gram de tesut, se produc aproximativ 2·1012 ionizari si 1013 activari fata de 2·1022 molecule de apa existente în constitutia a 1g de tesut viu. Astfel, se poate aprecia ca numai o molecula de apa dintr-un miliard este ionizata, neglijând ceilalti constituenti ai tesuturilor. Pentru a ioniza 50% din moleculele de apa prezente în 1g de tesut este necesara o iradiere cu o doza de 100 R/min timp de 200 de ani.
Radicalii liberi formati se pot recombina, formând:
Apa: H· + OH· H2O
Hidrogen molecular: H· + H· H2
Apa oxigenata: OH· + OH· H2O2
catalaza
Apa oxigenata
din celule se descompune rapid în apa si oxigen sub actiunea
unei enzime active, catalaza:
![]() H2O2 H2O + ½ O2
H2O2 H2O + ½ O2
c) actiunea la distanta
Prin fenomenul de transfer de energie actiunea finala a reactiei se desfasoara în alt loc al structurii biologice decât cel în care s-a produs absorbtia initiala de energie. Transferul energiei se realizeaza prin transmiterea intermoleculara si intramoleculara a energiei.
Efectele celulare
Cuanta de energie care patrunde în celula interactioneaza cu atomii si moleculele componente ale acesteia. Interactiunea fizica duce la aparitia unor procese radiochimice ce au ca rezultat modificarile însusirilor biochimice si apoi a celor biologice ale celulei.
Absorbtia de energie nu depinde de compozitia chimica a substantelor absorbante ci de cantitatea de substanta. Astfel, cea mai mare parte din energia incidenta va fi absorbita de componentele care ocupa cea mai mare parte din volumul celulei - apa. Se stie ca majoritatea organismelor vii au în compozitia lor un procent mare de apa. La om si la mamifere el ajunge, în anumite tesuturi (muschi, creier) la 85%. Astfel, energia radiatiei ionizante patrunse în organism este absorbita în principal de moleculele de apa.
Radioliza apei are efect biologic indirect datorita radicalilor liberi precum H· si OH·, H2O2 si alti oxizi organici, care se formeaza. Acestia, datorita potentialului chimic foarte mare, intra în reactie si modifica structura chimica a macromoleculelor esentiale pentru celule (proteine, acizi nucleici, enzime etc).
Macromoleculele pot fi supuse actiunii radiatiilor si în mod direct, dar posibilitatea ca ele sa absoarba direct energia radiatiei care patrunde în celula este mica.
Radicalii liberi formati pot determina afectarea atât a cromozomilor (moleculele de AND) cât si a proteinelor citoplasmatice. Modificarile enzimelor (directe sau indirecte) pot duce la aparitia unor metaboliti cu actiune mutagena determinând modificari genetice ce se transmit la descendenti.
Eficienta radicalilor liberi care apar prin radioliza apei este limitata la timpul scurt de viata a acestora. Ei traiesc numai 2·10-7 s, timp în care pot parcurge o distanta de 30-50 . Fata de celula ca întreg si chiar fata de spatiile intercelulare, aceasta distanta este relativ mica, ceea ce arata ca sfera de influenta a efectelor directe si indirecte este destul de limitata.
Prezenta oxigenului dizolvat în apa, duce la aparitia radicalului hidroxilic oxidat ·HO2, de asemenea, H2O2 si hidroxizi organici stabili. Compusii peroxidati au timp de viata mai lung si, astfel, sunt mai nocivi.
Efectivitatea biologica relativa (factorul de calitate Q) reprezinta raportul dintre energia absorbita de un tesut la o anumita doza de iradiere raportata la energia absorbita de acelasi tesut la iradierea cu aceeasi doza de radiatie standard (radiatie X de 200 KeV). Pentru radiatia X, g sau b, Q = 1, pentru neutronii termici Q = 5, pentru neutronii rapizi si protoni Q este 10 iar pentru a este 20.
Un organism viu, pus în variate conditii de mediu, poate reactiona diferit la aceeasi doza de iradiere. Reactia celulelor la iradiere depinde de proprietatile lor interne (grad de oxigenare, de hidratare, pH, grad de diferentiere), de existenta unor sisteme de aparare naturala (echipamente enzimatice implicate în refacerea structurilor lezate).
Pentru aprecierea efectelor radiatiilor ionizante, se folosesc mai multe marimi si unitati, prezentate în tabelul urmator:
MĂRIMI FOLOSITE ÎN RADIOBIOLOGIE sI MEDICINA NUCLEARĂ
|
Denumire si definitie |
Simbol |
Unitati S.I. |
Unitati tolerate |
Relatii de transformare |
|
RADIOACTIVITATEA (viteza dezintegrarii radioactive) |
L |
Bq Bequerel |
Ci Curie |
1Bq = 1dps = 27,027·10- 12 Ci 1mCi = 37 Bq 1Ci = 3,7·1010dps |
EXPUNEREA LA RADIAŢII(doza incidenta) D = q r V Numarul de sarcini electrice q produse în volumul V de aer, unde r este densitatea aerului |
D |
C/Kg Coulomb pe kg aer |
R (*) Roentgen |
1R=2,58·10-4 C/kg aer |
DOZA ABSORBITĂDabs = E/m Raportul dintre energia E, depozitata într un mediu oarecare si masa m a mediului |
Dabs |
Gy Gray |
rad |
1Gy=1J/Kg=100 rad 1 rad = 0,01J/Kg |
|
DOZA ECHIVALENTĂ BIOLOGIC Dbio = Q D Factorul de calitate Q, tine seama de faptul ca pentru doze absorbite egale, diferitele feluri de radiatii ionizante, sau acelasi fel de radiatii dar de energii diferite, produc efecte biologice diferite. |
Dbio |
Sv Sievert |
rem |
1Sv = 1J/Kg 1Sv= D(Gy)·Q·N = 100 rem 1rem=0,01Sv=10mSv |
* In conditii normale n aer, 1 R produce 2,1 x 109 perechi ion-electron pe cm3
|
