Calitatile motrice atletism
Calitatea motrica - in conceptia teoreticienilor reprezinta calitatea musculara determinata de factori biologici, morfologici si functionali dar si factori de ordin psihologic.
Atat biologii cat si psihologii definesc calitatea motrica ca fiind insusirea esentiala a activitatii musculare exprimata prin intermediul actelor motrice conditionata de structura morfologica si capacitatile functionale ale diferitelor aparate si sisteme ale organismului dar si de procese si capacitati psihice.
Argumentele necesare pentru a raspunde la intrebarea de ce 'calitati motrice' si nu 'calitati fizice' sunt date de dictionarul limbii romane contemporane care explica si se fac referiri la corpul fiintelor vii - in special la activitatea musculara. Astfel, conceptia notiunii de calitati fizice are un continut mult prea limitat privind numai determinarile de ordin biologic (respectiv morfologic, biochimic si functional) fara a lua in consideratie faptul ca activitatea motrica - ca orice forma a activitatii umane - are si determinari de ordin psihic: procese, capacitati si stari psihice.
Viteza, forta, rezistenta sunt calitati motrice de baza, iar indemanarea dupa opinia multor specialisti (Matveev, Harre, Novikov) nu poate fi admisa intre calitatile motrice de baza, ea fiind o calitate complexa, combinata, respectiv mai mult o capacitate motrica. In actul motric ele se manifesta in combinatie, in diferite proportii una avand rol preponderent. ( T. Ardelean, 1991).
Calitatile motrice se pot dezvolta peste nivelul atins in evolutia naturala a orgasnismului.
Studiul calitatilor motrice reprezinta un domeniu vast de aprofundare pentru specialistii domeniului miscarii, intrucat misscarea nu inseamna deplasare simpla, dezordonata; ea se realizeaza avand la baza functionalitatea organelor, transformarile energetice si procesele nervoase si psihice. Toate acestea se realizeaza in stransa legatura unele cu altele, influentandu-se reciproc. In literatura de specialitate apar insa denumiri si interpretari diferite, in functie de specialitatea teoreticianului (pedagog, psiholog, biochimist, fiziolog, teoretician sportiv). Gundlach, 1968, a propus o clasificare a aptitudinilor motrice care ulterior a fost acc 757f52h eptata si in toata Europa. Acesta imparte aptitudinile in doua categorii:
a) aptitudini conditionale: aptitudinea de viteza;
aptitudinea de rezistenta;
aptitudinea de forta.
b) aptitudini coordinative care sunt conditionate de capacitatea de a dirija si prelucra informatii provenind de la analizatorii implicati in miscare (tactil, optic, vestibular, chinestezic).
Zatiorschi considera ca notiunea de calitati fizice 'constituie premize sau procerinte motrice de baza, pe care omul, sportivul isi cladeste propriile abilitati tehnice'.
Renato Manno, 1992, in lucrarea 'Bazele antrenamentului sportiv' prezinta calitatile motrice sub denumirea de aptitudini motrice pe care le considera ca fiind 'de fapt premizele motrice de tip endogen, care permit formarea abilitatilor motrice'.
J. Weineck, 1990, prezinta principalele forme ale solicitarii fizice sub forma calitatilor fizice ca: anduranta, forta, viteza, mobilitate, coordonare.
T. Ardelean defineste calitatea motrica ca fiind 'aptitudinea individului de a executa miscari exprimate in parametri de viteza, de forta, de rezistenta, de indemanare'.
Calitatile motrice exista si se manifesta drept calitati ale actiunilor motrice. Ele sunt insusiri ale sistemului neuro-muscular de a produce miscari propriului sau corp, de a imprima miscari altor corpuri, respectiv obiectelor de aruncare (greutate, disc, sulita, ciocan) aparatelor si obiectelor ajutatoare in procesul de pregatire '.
In constitutia unei probe atletice, a unui exercitiu, calitatile motrice se gasesc intr-o anumita interrelatie, o anume combinatie. Pregatirea rezultatului sportiv depinde de gasirea acelei metodologii, care sa asigure raportul optim de dezvoltare specific probei sp practicata si particularitatile individului.
Viteza
Viteza este definita in literatura de specialitate in mod diferit, in conformitate cu conceptia si specialitatea celui care se ocupa de acest lucru.
M. Pradet, 1996, prezinta viteza ca fiind capacitatea de a executa o serie de actiuni motrice, provocand cea mai rapida deplasare a corpului sau a uneia din partile sale in unitati scurte de timp, fara sa duca la aparitia semnelor de oboseala.
La aceasta definitie, acelasi autor adauga urmatoarea explicatie: pentru a identifica exclusiv aceasta capacitate, este necesar, ca aceste actiuni sa nu fie obstacolate de niste rezistente externe importante, putandu-se manifesta in conditii in care sportivul nu trebuie sa infrunte decat efectele gravitatiei.
In conceptia profesorului L.Seagrave, 1996, viteza este capacitatea de a trece dincolo de limita si control; realitatea este ca nimic nu poate limita potentialul de imbunatatire al vitezei. Predispozitiile genetice vor influenta orice capacitate umana; cea mai mare greseala care se face in antrenamentul atletic este de a subestima potentialul care exista pentru fiecare atlet. Daca un atlet nu-si demonstreaza evident abilitatile ca sprinter la o varsta frageda sau de la primul antrenament, nu este obligatoriu ca acesta sa fie orientat catre alte probe sau discipline.
Acelasi autor defineste viteza de alergare, ca fiind si rezultatul tehnicii de alergare. Ca orice tehnica, acestea pot fi invatate si perfectionate, de catre cei care cunosc metodologia specifica.
In studiile lui N. Sozanscki si T. Witeczac, 1979, viteza este o calitate motrica, complexa, care este determinata de 3 componente principale:
* timpul de reactie;
* viteza unei miscari singulare;
* frecventa miscarilor.
Fiecare componenta este influentata genetic si are o serie de conditionari specifice, determinand astfel ca viteza in actiunile motrice complexe sa apara intr - o forma integrala. E. Hahn, 1996, denumeste viteza unei miscari singulare ca fiind viteza aciclica sau viteza de actiune, care este capacitatea de a executa o miscare in cel mai scurt timp posibil.
Frecventa miscarilor o denumeste viteza ciclica sau viteza de coordonare, ca fiind tot capacitatea de a executa miscari ciclice, in cel mai scurt timp posibil si a le incheia fara oboseala.
M. Pradet, 1996, considera ca intr-un prim stadiu, dezvoltarea unei calitati fizice determina progresul ansamblului celorlalte.
De exemplu, in fig. de mai jos, ameliorarea indicilor de rezistenta duce la dezvoltarea celor de indemanare, forta, viteza. Dupa acest stadiu, dezvoltarea nivelului unei anumite calitati nu pare sa mai exercite efecte pozitive asupra celorlalte calitati.
![]()

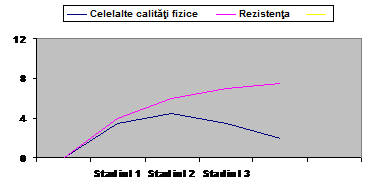
perioada de pregatire
Fig. nr. 2. Raportul dintre o calitate dezvoltata si celelalte calitati fizice, in functie de nivelul de experienta al sportivului. Dupa M. Pradet, 1996.
Din figura anterioara, observam ca viteza, ca si forta si indemanarea in cele 3 stadii de pregatire ale atletului, se afla fata de rezistenta pe o curba ascendenta in stadiul 1, apoi creste si atinge varful de dezvoltare spre sfarsitul stadiului 2, iar in stadiul 3, de pregatire, in timp ce rezistenta se dezvolta, viteza intra in perioada de regres.
Factorul genetic si capacitatea motrica viteza
In timp ce genetica determina care dintre noi are cele mai bogate resurse, nimic nu impiedica sa se exploreze si sa se descopere alte limite ale potentialului uman. Complexul genetic individual isi pune amprenta pe toata perioada performantei din viata unui sportiv. Din acest punct de vedere, capacitatea de viteza demonstreaza impactul structurii genetice asupra unui atlet.
In conceptia lui J. North, 1992, mecanismul neurologic de control al miscarii este determinat de structura ierarhica a sistemelor motorii centrale. Programul motor isi are locul in cortexul premotor, in aria motorie suplimentara si alte arii corticale asociate. Aceste arii, impreuna cu impulsul senzorial, converg catre aria motorie primara, care eventual excita sau inhiba partile specifice ale sistemului nervos. Programul motor este sistemul de analizare a informatiei esentiale care controleaza miscarile stereotipe. El comanda muschilor sa se contracte intr-o ordine prestabilita, la nivele de forta si timp determinate, pentru a produce miscari predeterminate (R. Enoka, 1994).
R.A. Schmidt, 1999, sustine ca un program motor este structura neurofiziologica de analiza (retea neurala), intarita de un model specific de succesiune a etapelor invatarii motorii.
Un alt aspect referitor la influenta zestrei ereditare privind viteza de alergare este efectul genetic asupra distributiei tipurilor de fibre si activitatii enzimelor in reactiile metabolice privind rolul de energetician a ATP-ului, CP-ului si glicogenului in muschiul scheletic Stiind ca viteza se bazeaza pe efort anaerob, J.A. Simoneau, G.Lortie, C.Leblanc si C. Bouchard, 1986, au aratat ca intr-un efort anaerob de scurta durata, (10s pe cicloergometru) 70% se bazeaza pe zestrea ereditara, cand datele au fost exprimate per Kg de masa musculara activa. Preponderenta tipurilor de fibre musculare va influenta abilitatea de a alerga cu viteza mare pe distanta scurta.
Privind raportul in procente dintre tipurile de fibre participante in efortul de viteza, J.H. Wilmore, D.L. Costill si M. Rieu, 1986, sustin ca, un sprinter are 60-65% fibre rapide si 35-40% fibre lente. J. H. Wilmore si D.L. Costill, 1994, au raportari de tip fiziologic si mai recente despre efectul genetic asupra distributiei tipurilor de fibre si activitatii enzimelor in muschiul scheletic. Din acest punct de vedere, viteza este o incarcare dinamica de o intensitate maxima cu caracter aproape exclusiv anaerob. Ea este un efect al spasmelor energetice si musculare. Acest factor este ereditar si se bazeaza pe valoarea incarcarii celulelor ganglionare. De aceste spasme scurte si rapide va depinde viteza alergatorului.
Un alt aspect privind caracterul ereditar al vitezei apartinand tot autorilor de mai sus, este ca, in filamentele de actina exista 3 tipuri de proteine care intervin in controlul activitatii ATP-azice a miozinei: actina, tropomiozina si troponima care sunt de fapt proteine reglatorii privind ATP-ul, materia prima in efortul de viteza maxima.
C. Capizzi si altii, 1978, citat de Manno, 1992, au demonstrat ca subiectii rapizi au o viteza mai mare de conductie a influxului nervos, calitate care este de ordin genetic, respectiv constitutional. Aceste date, considerate global, sugereaza ca activitatea anaeroba de scurta durata este destul de mult influentata de genotipul indivizilor. In acest moment nu se cunosc date sigure privind ereditatea si lucrul anaerob de lunga durata.
Vom prezenta in tabelul de mai jos, o parte a studiului lui E.L. Fox si D.H. Mathews, 1984, care au demonstrat in ce masura unele functii ale organismului sunt ereditare:
|
Functii |
Sex |
Influenta ereditatii % |
|
Frecventa cardiaca maxima |
M | |
|
Tip de fibra |
M | |
|
F | ||
Putere mecanica |
M | |
|
F |
n.s. |
|
|
M+F | ||
|
Concentratia maxima de acid lactic |
M | |
|
Forta |
M+F |
n.s. |
|
Timp reflex al rotulei |
M | |
|
F |
n.s. |
|
|
Timp de reactie |
M | |
|
F |
n.s |
|
|
Viteza de conducere a fibrelor nervoase |
M+F |
n.s |
n.s.- componenta genetica nesemnificativa
Tabel nr.2.
In opinia fiziologilor Demeter, 1983, M. Malina si C. Bouchard, 1991, viteza de alergare in probele de sprint depinde in cea mai mare parte de principalii factori ereditari:
-viteza de contractie a muschiului este determinata de aparitia excitatiei nervoase viteza de contractie a muschiului depinde de timpul de inervatie de la nivelul placii motorii;
-viteza de conducere al influxului nervos aferent si eferent de-a lungul relatiei nervoase; viteza este rezultatul actiunii motoneuronilor alfa mari cu originea in coarnele anterioare ale maduvei spinarii, ce transmit inpulsuri nervoase rapide si discontinue prin mielina axonica; -mobilitatea desfasurarii proceselor nervoase: viteza alternarii proceselor de excitatie cu cele de inhibitie dar si intensitatea cu care acestea se manifesta.
Studiul lui J.P. Eclache, 1988, releva numeroase aspecte privind impulsul nervos:
* mesajele nervoase iau nastere in ariile cortico - motrice specifice ale fiecarui muschi sau grup muscular. Experienta si invatarea, mai cu seama in copilarie si adolescenta, dupa cum am mentionat deja se afla la originea dezvoltarii si consolidarii legaturilor neuronale; se formeaza noi proeminente dendritice, inmultindu-li-se numarul, iar terminatiile presinaptice se largesc; celulele neuronale sintetizeaza ele insele proteine specifice membranoase, care asigura conservarea mnemonica a mesajelor anterioare repetate si le sporesc posibilitatile de sinteza si de eliberare a transmitatorilor.
Aceasta programare centrala permite atunci, pornindu-se de la un singur ordin motor, intrarea in actiune a circuitelor preferentiale caracteristice de scheme gestice deosebit de bogate si complexe.
* bogatia legaturilor intercentrale ingaduie modularea acestei programari prin intermediul stimularilor excitatoare sau inhibitoare provenind de la captatorii sensibili la modificarile fizico - chimice ale mediului exterior sau interior. Aceasta modulare exista la toate nivelele sistemului nervos central si periferic.
* toate stimularile mai vii (auditive, vizuale, tactile), care exercita un efect excitant asupra retelei fibrelor nervoase cerebrale, sunt insotite de o reactie de punere in garda, de ascutire a atentiei si a comenzii motrice (J.P. Eclache si M. Jaunet, 1986). Posibilitatile asemanatoare apar si in controlul spinal: intinderea manunchiurilor neuromusculare (bucla gamma) exercita un efect excitant asupra comenzii motrice; invers, stimularea organelor Golgi si a celulelor Renshaw exercita un efect inhibitor .
* in sfarsit, trebuie sa tinem seama de faptul ca transmiterea sinaptica, excitabilitatea, ca si conductabilitatea nervoasa sunt susceptibile a fi modificate prin caracteristicile fizico chimice ale mediului interior; daca acestea sunt relativ stabile, acest fapt se datoreaza protectiei pe care o asigura bariera hematoencefalica la nivel central. In schimb, temperatura, cresterea procentului de catecolamine, de amfetamine etc. pot exercita un efect excitant la nivel periferic.
Viteza, capacitate motrica potentiala
In conceptia lui T. Ardelean, 1991, viteza este o calitate potentiala iar valorificarea potentialului de viteza depinde de maiestria practico-metodica a fiecarui profesor de specialitate. N. Sozanscki, 1979, citat de T. Ardelean, 1991, defineste viteza ca fiind nivelul potentialului motric si al pregatirii specifice.
Sub pretextul ca, la copii, trebuie sa se pastreze intact potentialul de evolutie a calitatii de viteza, pentru a fi valorificat corect, atat sportivii cat si antrenorii ajung uneori la deziluzii, cand, fata de copil, se manifesta din partea adultilor, pe de o parte un ,,hiperprotectionism caruia i se interzice orice efort susceptibil pentru dezvoltarea fizica si, pe de alta parte, o ,,tehnicizare excesiv de timpurie.
Pentru a nu se ajunge la aceste aspecte negative in pregatirea copilului, potentialul de viteza trebuie valorificat prin a asigura atletului incepator un nivel de instruire tehnica in paralel cu abordarea vitezei in probele de alergari, apoi in probele de sarituri si aruncari si, de asemenea, de a participa la pregatirea de forta, care este conditionata de varsta si nivelul de insusire a unui grup de probe asemanatoare, evitand pe cat posibil instalarea barierei de viteza la varste fragede, dar si mai tarziu. (Ardelean, 1991)
Specialistii francezi M. Pradet, 1996, A. Duray, 1997, au concluzionat ca, pentru a valorifica, prin tehnica, potentialul de viteza, trebuie precizate si apoi respectate anumite principii de lucru, valabile la toate nivelurile de dezvoltare a vitezei:
- pentru a imbunatati viteza unui element tehnic este necesar ca acesta sa fie repetat de multe ori, dar cu cat elementul se repeta mai mult, cu atat riscul de a ajunge la o stagnare in dezvoltarea vitezei este mai mare, ajungandu-se la instalarea barierei de viteza.
- metodele de actionare analitica impotriva acestei bariere se vor utiliza indeosebi cu atletii care au depasit pubertatea, iar in plan tehnic trebuie sa se actioneze astfel incat, tehnica miscarii sa nu fie alterata de variatiile rezistentelor.
Studiile lui G. Dintiman, 1997, L. Seagrave, 1996, C. Vittori, 1990, sustin ca la fel se petrece si cu capacitatea de accelerare, care e relativ independenta de capacitatea de a alerga cu viteza maxima.
Concluzia autorilor de mai sus este ca, pentru a fi sprinter cu o tehnica eficienta de alergare este nevoie a se respecta urmatoarele aspecte:
*executia unui element tehnic cu viteza lenta nu este transferabila la viteze apropiate de maxim;
*transferul de viteza nu se efectueaza decat in miscarile in care coordonarile sunt asemanatoare.
Sa nu uitam ca succesele precoce sunt de multe ori sanctionate mai tarziu, intr-o viata sportiva efemera.
Viteza, rezultat al procesului metabolic
Muschiul reprezinta motorul miscarii umane si necesita o anumita sursa de energie. Acesta se prezinta sub forma unor molecule bogate in elemente fosfor; ATP adenozin trifosfat, care se afla in muschi in cantitati de 6mMol/kg muschi umed pentru un individ cu greutatea de 70kg, iar copiii poseda ATP cu 30% mai putin decat adultii, dupa cercetarile fiziologilor M.R. Malina si C. Bouchard, 1991.
Atunci cand fibrele musculare sunt stimulate de influxul nervos, aceasta molecula de ATP, situata la nivelul acelor fibre, prezinta particularitatea de a se disocia, eliberand energia in masura sa produca contractia lor cu intensitate maxima.
Acest fapt permite efectuarea unor exercitii foarte intense, cu o putere maxima, ceea ce se dovedeste extrem de util in alergarile de viteza. In schimb, acest mecanism nu are puterea sa intretina mult timp contractia musculara. Fiind solicitat la maximum de intensitate, trebuie sa se tina cont de faptul ca el se epuizeaza dupa circa 6 secunde. Acest timp, de 6 secunde, dupa unii fiziologi europeni, iar dupa cei americani, de 3-4 secunde, reprezinta un reper esential in elaborarea mijloacelor de dezvoltare a acestui proces. Aceste aspecte releva ideea, ca, existenta sistemului anaerob se raporteaza la o capacitatea mica.
Reactiile chimice care stau la baza resintezei ATP consuma creatin fosfatul in cantitate mare. In absenta unei rezerve energetice, miscarea din timpul antrenamentului trebuie sa stabilizeze concentratii superioare de ATP si CP intramusculare si o utilizare mai eficienta a energiei produse. Acest aspect se pare ca este aprofundat de C.G. Stathis si col., 1994, exemplificand rezervele de fosfogeni numai intr-un singur muschi vastus lateralis.
Respectivii autori analizeaza influenta antrenamentului de sprint asupra metabolismului fibrei striate, mai exact cat se modifica cantitatea de ATP, CP, creatina, (mMol/kg. d.wt) aflate in muschiul vastus lateralis inainte, in timpul si dupa sprint, atat la subiectii antrenati cat si neantrenati, in varsta de 22,1 ± 1,0 ani:
ATP CP CREATINA
inainte de sprint:
neantrenati 22,8 85,4 42,9
antrenati 18,5 92,2 40,9
in timpul sprintului:
neantrenati 13,6 27,6 100,7
antrenati 14,1 29,6 103,3
dupa 3 minute de pauza:
neantrenati 16,3 65,5 62,8
antrenati 16,0 71,2 60,3
La investigarea initiala, cantitatea de ATP este mai mare inainte de sprint, in acest muschi, la neantrenati si respectiv, la antrenati mai mica. In timpul sprintului, antrenatii consuma ATP mai putin (4,4 mMol/Kg. d. w) in timp ce neantrenatii consuma o cantitate mai mult decat dubla de ATP (9,2mMol/Kg dry/wet). Dupa efort, cantitatea de ATP, la antrenati, ca valoare este mai aproape de nivelul initial, (respectand proportia) decat la neantrenati. Cantitatea de CP inainte de sprint este mai mare cu 6,8 mMol/Kg d.wt fata de neantrenati. In timpul sprintului, CP se consuma aproximativ la fel; dupa sprint cei antrenati au o cantitate mai mare de CP in respectivul muschi. Cantitatea de creatina ramane aprox. neschimbata in toate situatiile. Din acest studiu rezulta ca cel antrenat, in timpul sprintului consuma de 2 ori mai putin ATP decat neantrenatii.
G.C. Gaitanos si col., 1993, au observat ca dupa 10 repetari de 6s, cu pauze de 30s, ATP-ul din acelasi muschi de mai sus, a scazut de la 24 la 20,9mmol/kg(d.wt). Aceasta degradare neta a ATP-lui a fost apreciata ca fiind de 6% din productia totala de energie. Anterior celei de-a zecea repetare, nivelul ATP-lui a fost de 16,4mmol/kg(d.wt).
Intr-un alt studiu al aceluiasi autor, subiectii au executat 15 repetari de viteza timp de 10s, de doua ori pe zi, timp de 7 zile. Nivelul ATP-ului de repaus a fost mai mic decat cel normal: 16 mmol/kg (d.wt). Aceasta pierdere aparenta de ATP din muschi, a fost atribuita unei cresteri in fluxul reactiei de adenadine kinase, ca o consecinta a cererilor energetice extrem de mari, din timpul perioadelor de efort. Oricum, implicatiile, daca sunt, in reducerea marimii baltii de adeninnucleotide in muschii scheletici, in repaos, nu sunt deocamdata clarificate.
L.H. Boobis si altii, 1982, sustin ideea ca majoritatea nevoilor de ATP in activitatea contractila din timpul unei singure repetari, de 6s, este sustinuta anaerobic de degradarea creatinfosfatului si glicogenului. E. Shroubridge si G.T. Radda, 1987, considera ca reactia creatinkinasei este cea care debuteaza prima dintre aceste doua sisteme energetice si a fost prezentata ca fiind activata in mai putin de 200 milisecunde de la debutul contractiei musculare. Contributia energetica a creatinkinasei in timpul efortului rapid de 6s, a fost estimata la 50% din totalul de energie anaeroba. In timpul celei de a zecea repetare de viteza, de 6s, degradarea creatinfosfatului a fost estimata la o cantitate de 80% din totalul productiei energetice pe cale anaeroba.
Aceste experimente demonstreaza rolul important al CP-ului in timpul efortului maxim, de 6 s. J.M. Crielaard, 1981, a demonstrat puterea anaeroba alactacida, determinata pe o bicicleta ergometrica, fiind examinate 160 de fete in varsta de 12-18 ani. S-a dovedit ca la varsta de 12-14 ani, 28 de fete (care nu sunt sportive) din cele 160, care au participat la experiment, au PAA de 313,5W iar PAA relativa este de 6,48 W.Kg-1. De asemenea, masurandu-se PAA la fete aflate in perioada pubertatii, acestea au realizat valoarea de 370,2 W, iar fetele nepubere au obtinut o PAA de 264,4 W. Sportivele de aceeasi varsta au PAA cu 15-25% mai mare fata de nesportive.
In opinia lui M. Rieu, 1986, la varsta de 10 ani, fetele au o putere anaeroba de 60 J.Kg-1 in testul de 10 s pe bicicleta ergonomica si de 75 J.Kg-1 la varsta de 14 ani, putere ce se mentine pana la varsta de 18 ani, ca apoi sa creasca spre valori de 80J.Kg-1.
R. Malina si C. Bouchard, 1991, considera ca fetele si baietii in varsta de 10 ani au puteri egale si valoarea ei reprezinta jumatate din valoarea adultilor, iar la varsta de 14 ani, energia lor este de 90% din valoarea adultilor.
In acest sens, Van Schenau, 1994, prezinta cantitatea de energie consumata in fiecare faza a alergarii de sprint de atletii consacrati. In primele 4 -5 de alergare in proba de 100 mp, energia este sustinuta de o mare cantitate de fosfati cu un echivalent mecanic de 20W/Kg si chiar mai mult si apoi de 10W/Kg pentru a mentine viteza acumulata.
Astfel, un sprinter de 80 Kg care atinge viteza maxima de 11,5 m/s in primele 4, necesita o energie la start de 1092W.
La o viteza de 10,5 m/s, necesarul de energie este de 1050W iar la 9,5 m/s puterea este de 990W. Dar tot acelasi autor sustine ca in cursa de 100 mp, prin frecarea cu aerul se consuma o putere de 440W, pentru a mentine o viteza maxima de 11,5 m/s.
In faza de accelerare a unei curse de sprint de 100 mp este nevoie de o putere relativa de 20W/Kg la o viteza de 10 m/s.
La o viteza de 3,6 m/s specifica copiilor cu acceleratie de 10-11 m.s-2, energia ajunge pana la 3000W, ceea ce inseamna mai mult de 4 cai putere.
Concluziile acestui studiu sunt urmatoarele:
- puterea mecanica depinde de datoria de oxigen in timpul alergarii la diferite viteze;
- datoria de oxigen creste liniar cu viteza de alergare, in opinia lui Boileau et al. 1982, citati de Schenau 1994.
Prin comparatie cu cele afirmate mai sus, intr-un studiu realizat de A. J. Ward Smith si P. F. Radford, 2000, in cursa de 100 mp cu sportivi consacrati, s-a reliefat ideea ca diferentele de viteza maxima si distanta unde a fost atinsa se datoreaza modurilor diferite in care sexele transforma energia in timpul sprintului.
Consumul de energie al atletelor consacrate este de 492 J.Kg-1 si 475 J.Kg-1 la atletii de elita. 90,5% este energie anaeroba si 9,5% energie aeroba la atlete si 92,5% energie anaeroba si 7,5% este energie aeroba pentru atleti.
Declinul de viteza in cursele de sprint ale atletelor pe ultima parte a cursei este explicat prin declinul in sursele de energie anaeroba.
Metabolismul aerob joaca un rol semnificativ mai mare la sprintere decat la sprinteri, sustine acelasi autor. Astfel, se sugereaza ideea ca antrenamentul sprinterelor nu ar trebui sa-l imite pe cel al sprinterilor.
Metabolismul aerob este mai important pentru atlete de-a lungul tuturor distantelor de alergare. J. Ward Smith si P. F. Radfort, 2000, au observat ca atletele au folosit procente mai mari din rezerva aeroba decat atletii, in probele de sprint prelungit.
Tot J.Ward Smith si P.F. Radfort, 2000, si-au extins studiul si asupra puterii lactacide, demonstrand ca puterea relativa anaeroba lactacida, prin folosirea A.T.P.-ului endogen si C.P. la timpul de 21 in cursa de 100 mp, la Campionatul Mondial de la Roma, 1987, a fost de 45,8 WKg-1 la atlete.
In alta ordine de idei privind metabolismul contractiei musculare, se afla studiile care strabat in profunzime structura fibrei striate pentru a demonstra mai fidel si rolul microelementelor contractile in timpul contractiei excentrice.
In acest sens, C.M. Waterman - Storer, 1991, prin tehnici histochimice sofisticate, au pus in evidenta un alt tip de citoschelet exosarcomeric, care consta intr-un sistem de filamente intermediare, plasat in spatiul extramiofibrilar, dar tot in interiorul celulei musculare.
Ele se numesc desmina, vimentina si synemina si au rol in hipertrofia musculara de alungire, cand discul Z poate fi despicat. In citoscheletul endosarcomeric se gaseste un filament special, numit conectina, care este formata din titina si nebulina.
Ultimele doua au rol in capacitatea fibrei de a redobandi lungimea si functia contractila dupa supraintinderea muschiului, cand actina si miozina nu mai prezinta nici cea mai mica zona de suprapunere. Rolul acestor filamente este evident pentru contractia excentrica, care este atat de utila in efortul de viteza.
Nu exista nici o indicatie asupra rolului jucat de genotip in distributia tipurilor de fibre. Influenta genetica a enzimelor in acest sens este in proportie de 30-50% a fosfofructokinazei si 70% a lactat dehidrogenazei.
Viteza, rezultat al interferentelor factorilor psihici
Pornind de la teoria lui Hull, 1943, citat de R.S. Weinberg, 1979, performanta, care este produsul dintre forta deprinderii si motivatie, procesele cognitive si cele afective conduc la realizarea unor performante ridicate.
V.G. Payne,1980, citat de J.J. Temprado, 1989, foloseste termenul de comportamente decizionale contingente pentru a exprima ideea ca procesele cognitive nu sunt invariabile, ci depind de tratamentul informatiilor propriei sarcini.
F. Belinchon, 1981, sustine ca la obtinerea de performante sportive, sofrologia sportiva, ca metoda de integrare a personalitatii, are un rol important. Concurentii, care au deja o performanta, sunt mult mai constienti decat atunci cand participa prima data la concurs. Corpul face ceea ce ii spunem sa faca, daca il invatam sa se exprime, afirma acelasi autor.
Exista o serie de studii care demonstreaza ca vointa de a atinge viteza maxima este determinata atat de capacitati psihice bine conturate in conditii de stress, cat si de procese psihice implicate in efortul de viteza. Vom incerca in continuare sa punctam cateva dintre aceste aspecte.
1.Procese psihice implicate in efortul de viteza
F.C. Bakker, M.S. Boschker, T. Chung, 1996, au studiat efectele interferentei retroactive ale vitezei de miscare, imaginate mental. Intelegerea si constiinta de sine, ca procese psihice, au ca punct de pornire imaginea de sine. Aceasta imagine este alcatuita din toate experientele trecutului nostru, de la nastere sau de la debutul ca sportiv pana la momentul actual. Studiul lui F.C. Bakker, 1996, a avut drept scop sa descopere efectele vitezei unei miscari imaginate asupra tempoului miscarii efectuate in timpul unui efort fizic. Imaginandu-ti o actiune motrica la o anumita viteza, actiunea segmentului va fi influentata de viteza cu care ti-ai imaginat miscarea respectiva.
Fenomenul este cunoscut sub denumirea de tendinta de asimilare, fiind in acelasi timp si un raspuns al studiilor efectuate pana atunci, privind aceasta problema a miscarii imaginate. Annett, 1995 si A. Paivio, 1991, au demonstrat ca nu s-a gasit nici un efect aditiv al reperarii verbale asupra imaginarii vitezei miscarii.
Dar un atlet trebuie sa fie foarte atent ce-si imagineaza, pentru ca imaginile nepotrivite scopului alergarii ar putea produce diminuari insemnate ale performantei, releva J.J. Janssen si A.A. Sheikh, 1994.
R. A. Schmidt, 1991, sustine ca daca se ajunge la un automatism al prelucrarii informatiilor, multe actiuni se pot produce nu doar repede, dar si in paralel cu alte activitati, chiar fara a distruge performanta.
2.Capacitati psihice implicate in efortul de viteza
A. Zani si B. Rosi, 1991, considera ca metoda potrivita pentru cercetarea proceselor mentale in timpul eforturilor fizice si deci si in timpul eforturile de viteza este psiho-fiziologia cognitiva, prin care procesele mentale nu trebuie studiate separat de creier, acolo unde apar doua tipuri de potentiale. Astfel, reprezentarea stimulilor in sistemul nervos central se datoreaza potentialelor evocate (PE). Potentialele corelate cu evenimentele (PCE) sunt potentialele care se afla in corelatie cu succesele unor evenimente cognitive ca: orientarea atentiei, prelucrarea informatiei, reordonarea memoriei pe termen scurt pe baza rezultatelor prelucrarii, decizia pentru o eventuala executie a unei actiuni de miscare, sau de pregatire a actiunii. Metoda PCE a fost folosita de autorii de mai sus pentru studierea proceselor cognitive si de atentie la copii si adolescenti in varsta de 10,13,18 ani. Pentru a studia interactiunea procesului de maturizare a creierului la dezvoltarea atentiei, potentialele creierului au fost deduse din zonele partiale, centrale si frontale.
Autorii mai sus mentionati au concluzionat urmatoarele:
1*zonele frontale, care participa mai mult la procesele de atentie, ating maturitatea abia la varsta de 12-13 ani;
2*copiii in varsta de 10 ani au dovedit ca sunt mai putin eficienti la treapta de orientare a atentiei, prin care se intelege de fapt indreptarea rezervelor de atentie spre informatia in curs de receptionare, independent de relevanta acesteia pentru actiunea respectiva;
3*toti copiii in varsta de 10 ani, in ceea ce priveste aptitudinea de a deosebi informatiile relevante pentru o anumita problema, de cele nerelevante, sunt la fel de apti ca si adultii. Acest lucru s-a constatat si in ceea ce priveste timpii de reactie, la care copiii de 10 ani reuseau sa dea raspunsuri motorii foarte rapide la problemele relevante.
Aceasta concluzie confirma ipoteza ca performanta mai slaba in ceea ce priveste atentia la copii se datoreaza in mai mare masura inhibitiei reactiei motorii. Ambitia, motivatia sunt indispensabile in viata de sprinter, cu conditia ca ele sa fie in concordanta cu ceilalti factori esentiali, care, in conceptia lui D. Swartz, 1980, sunt:
-conditia fizica adecvata;
-tehnica corespunzatoare;
-igiena de viata (somn, alimentatie).
Respectand acesti factori, barierele mentale care limiteaza performanta fizica pot fi invinse. Acelasi autor sustine ca succesul in competitie se datoreste constientizarii de sine, prin imaginea de sine in postura victorioasa si a sentimentului valorii personale. Aceste capacitati psihice pot fi puse in evidenta prin vizualizare-concentrare a atentiei asupra unui eveniment viitor aflat in subconstientul nostru.
Desi stiinta a demonstrat ca subconstientul nu face diferenta intre evenimentele reale si cele imaginate, vizualizarea trebuie sa se faca la persoana I si in ajunul cursei: alerg inalt, accelerez si, in final de cursa, ma desprind de departe.
|
