Cuantificarea metabolismului energetic la efortului fizic
Abstract Estimarea productiei si consumului de energie (metabolismul energetic) in timpul efortului fizic presupune determinarea energiei produse atat aerob cat si anaerob. Determinarea consumului de oxigen atat prin masurarea acestuia la nivelul plamanilor cat si prin inmultirea oxigenului extras din sange cu viteza the curgere a acestuia, este un bun indicator al productiei aerobe. Productia anaerobica de energie este in schimb mult mai dificil de masurat la nivelul intregului organism, dar poate fi destul de precis estimata in exercitiile care angreneaza grupe musculare izolate. Deficitul de oxigen poate fi folosit in aprecierea productiei anaerobe de energie in cadrul exercitiilor the intensitate submaximala si probabil in exersarea folosind grupe musculare izolate in timp ce pentru exercitiile maximale folosirea acestuia poate da valori gresite in ceea ce priveste productia anaerobe de energie. Masurarea debitului de oxigen nu poate fi folosita in determinarea contributiei energeticii anaerobe la productia totala de energie in timpul eforului fizic.
Cuvinte cheie: metabolism energetic, energetica, aerob, anaerob, deficit de oxigen, debit de oxigen
Nu este o sarcina usoara sa investighezi viteza de producere si de consum a
energiei in timpul efortului si de asemenea diferitele surse din care energia
este furnizata pentru sustinerea acestuia. Dificultatea vine de la
posibilitatile limitate de masurare precisa a producerii si consumului de
energie in corpul omenesc in timpul exercitiului fizic.
Metabolismul energetic in muschi poate fi determinat fie printr-o abordare metabolica sau calorimetrica (Woledge 1998). Abordarea metabolica presupune insumarea energeticii aerobe si anaerobe, in timp ce calea calorimetrica consta in masurarea productiei de caldura a muschiului si rezistenta exteri 141h78b oara, incarcatura lucrului mecanic efectuat (i.e. rezistenta externa care determina intensitatea contractiei). In ce priveste abordarea metabolica, estimarea atat a diviziei aerobice cat si a celei anaerobice ale productiei totale de energie, poate fi facuta atat prin masurari directe cat si indirecte.
Productia aeroba de energie
Productia aeroba de energie se termina cand molecula de oxigen accepta doi atomi de hidrogen si formeaza apa in cadrul lantului respirator din mitocondrii. Absorbtia de oxigen poate fi masurata atat la nivelul plamanilor cat si prin multiplicarea oxigenului extras din sange in timpul exercitiului cu viteza de circulatie a acestuia. Consumul de oxigen determinat la nivelul plamanilor preeupune masurarea cantitatii de oxigen din aerul respirat in timpul ventilatiei pulmonare. Aceasta poate fi facuta fie prin colectarea aerului expirat in saci special cum ar fi de exemplu sacii Douglas, sau prin folosire unui sistem on-line computerizat care efectueaza masuratorile automat, furnizand informatiile in timpul efectuarii exercitiului.
Pe langa tehniciile si masuratorile efectuate in investigarea consumului de oxigen in timpul efortului, este de asemenea foarte important de evaluat si relatia dintre absorbtia oxigenului si lucrul mecanic efectuat. In urma cercetarilor efectuate in studii avand ca subiecti oameni, s-a descoperit ca in cadrul exercitiilor de intensitate submaximala, aceasta relatie este liniara, in timp ce la intensitati submaximale mari si maximale, relatia dintre consumul de oxigen si incarcatura exercitiului devine curviliniara (Bangsbo et al 1993; Zolads et al 1995; Pedersen 2002). Aceasta relatie non-liniara are implicatii in discutarea energeticii efortului fizic si eficientei musculare, cat si in ceea ce priveste testarea indirecta a consumului maxim de oxigen in timpul efortului sau a deficitului acestuia in perioada de recuperare post efort, care va fi discutata mai tarziu in cadrul acestui articol. Explicatia cea mai des folosita in legatura cu relatia consum de oxigen-incarcatura efortului, este aceea ca modul de recrutare a fibrelor musculare in muschii angajati in efort, se schimba de la cele care oxidative (tipul I) care sunt recrutate in special la intensitati scazute al efortului, catre cele glicolitice (tipul II) care sunt predominant recrutate la intensitati mari si viteze mari de contractie (Gollnick et al 1974; Vøllestad et al 1985). Fibrele de tipul II au fost descoperite a avea o eficienta mai scazuta din punctul de vedere al folosirii oxigenului in producerea aeroba de energie pentru aceasi intensitate a exercitiului in comparatie cu fibrele de tipul I (Crow et al 1974; Barclaz et al 1993). In aceasta privinta, P/O (productia de ATP/consumul de oxigen) s-a descoperit a fi mai scazuta in fibrele glicolitice (Willis and Jackman 1994) si aceasta este reflectata in consumul de oxigen crescut de la nivelul plamanilor.
Oxigenul consumat in timpul efortului poate fi convertit in kilocalorii sau kilojouli (kcal sau kJ). In tabelul 1 se poate observa echivalentul energetic al oxigenului consumat exprimat de coeficientul respirator (RQ) non-proteic. Totusi, se impun cateva considerente. Non-proteic RQ presupune ca in timpul exercitiului carbohidratii si grasimile sunt sursele de energie care predomina. Mai mult, pentru ca RQ reprezinta raportul dintre dioxidul de carbon produs si oxigenul consumat (CO2/O2) este important ca acesta sa fie masurat la nivelul celulei musculare. Aceasta pentru ca, in timpul exercitiilor de intensitate mare lactatul eliberat de muschi reactioneaza cu bicarbonatul din sange, reactie chimica care se termina cu formarea de CO2 si H2O. Prin urmare are loc o crestere aditionala a CO2 din sange, care in final va fi descarcat in aerul alveolar. Calcularea RQ folosind acest supraestimat CO2 nu va reflecta adevarata CO2/O2 de la nivelul celular. Privind mai atent la tabelul 1, se poate constata ca in jumatatea de sus a coloanei care prezinta RQ corespunzator oxigenului consumat, valorile acestuia sunt mai scazute, indicand ca sursa predominant folosita in producerea energiei este reprezentata de grasimi si de carbohidratii oxidati aerob. Odata cu cresterea intensitatii exercitiului, sursa de energie folosita predominant o formeaza carbohidratii care, cand sunt in intregime oxidati aerob in cadrul intensitatilor care nu implica formarea de lactat, dau un raport al RQ-ului egal cu 1.00. In cadrul eforturilor de intesitati mai mari, glucoza este in primul rand si intr-o mai mare masura utilizata in cadrul glicolzei anaerobe pentru producerea ATP-ului, cu formare de acid lactic. Cu toate acestea in cadrul exercitiilor care implica grupe musculare izolate, directa estimare a productiei anaerobe de energie prin determinarea schimbarilor in metabolitii din muschi, cum ar fi ATP, fosfocreatina (FCr) si acumularea de lactat in muschi si eliminarea lui in sange in timpul efortului, face ca determinarea producerii aerobice de energie sa fie mai precisa. Pentru aceasta insa, trebuie recoltate biopsii musculare si asta ar putea fi un incovenient in sportul de inalta performanta, unde programele de pregatire elaborate in vederea imbunatatirii si mentinerii formei sportive, nu permit lungi periode de timp pentru recuperarea muschilor biopsiati in urma acesui fel de interventi.
|
Non-protein RQ |
kcal L-1 O2 |
kJ L-1O2 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
Tabel 1. Echivalentul
energetic pentru
Relatia dintre absorbtia de oxigen si lucrul mecanic efectuat, a fost cel mai des studiata, la o intensitate a efortului care a fost asociata cu aparitia pragului de lactat, concentratie la care se constata o usoara acumulare de lactat in sange. Aceasta denumire semnifica cea mai mare valoare a absorbtiei de oxigen la care concentratia de lactat din sange ajunge la 1 milimol per litru, peste nivelul lactatului din sange din perioada de repaus. Inceputul de acumulare a lactatului in sange este reprezentat de o valoare a lactatului de 4 milimoli per litru de sange. Acesta, ca si pragul de lactat, este reflectat in intensificarea disproportionata a ventilatiei pulmonare, proces care semnifica pragul ventilator. In studierea relatiei dintre distributia fibrelor musculare in muschi si relatia non-liniara dintre consumul de oxigen si lucrul mecanic efectuat in timpul pedalarii pe bicicleta stationara, Pedersen et al (2003) au stabilit ca si criterii pentru efectuarea unui efort maximal, realizarea a cel putin doua din urmatoarele performante: 1) Rata schibului repirator (masurata la nivelul plamanilor) ≥ 1.10; 2) lactatul din sange ≥8.0mmol L-1; 3) echivalentul in ventilatie pulmonara pentru consumul unui litru de oxigen ≥30 L L-1. O alta optiune poate fi stabilirea unei intensitati a exercitiului care sa se plaseze la jumatatea distantei (50%) dintre pragul ventilator si consumul maxim de oxigen in timpul exercitiului (Pringle et al 2003). Masurarea acestor parametri are loc in experimente preliminarii studiului principal.
Estimarea absorbtiei oxigenului prin masurarea extractiei oxigenului din sange si multiplicarea lui cu viteza de curgere a sangelui, este mult mai precisa dar numai in exercitiile care implica un numar limitat de grupe musculare si rezultatele nu pot fi raportate la intregul organism.
Productia anaerobe de energie
Aceasta poate fi determinata atat prin masurarea deficitului si debitului de oxigen, metode care reprezinta o abordare indirecta, cat si prin masurarea directa a degradarii ATP-ului si FCr si acumularea de lactat in muschii angajati in efort si eliminarea acestuia in circulatia sangvina. In ceea ce priveste a doua cale, cea directa, o imagine complexa dar si clara este data de formula utilizata in studiul lui Bangsbo et al (2001), in care ei au masurat contributia anaerobica la productia totale de energie folosind urmatoarea formula: ΔATP + ΔCP + 3/2Δ muscle lactate + 3/2 (lactate si pyruvate release) + "altii", unde ATP reprezinta cantitatea neta de ATP consumat, presupunand ca primele schimbari in concentratia de ATP din muschi se observa dupa 15 secunde (Hellsten et al 1999). "Altii" reprezinta productia de ATP corespunzatoare acumularii de acid piruvic, care este estimata a fi 1/30 din lactatul acumulat in muschi (Spriet et al 1987), cantitatea de lactat absorbita de fibrele si/sau grupele musculare inactive din muschii angajati in efort (Bangsbo et al 1995), acumularea de intermediari in lantul de reactii chimice care formeaza glicoliza (Spriet et al 1987) si acumularea si eliminarea aminoacidului alanina (Katz et al 1985). Inca odata, acest fel de investigatie poate fi facut doar pe o parte restransa a corpului omenesc care poate include una sau mai multe grupe musculare.
Unul dintre cele mai folosite modele de exercitiu in fiziologia sportiva este extensia piciorului din asezat (Fig 1). Aceasta pentru ca, bazat pe masuratori facute prin folosirea EMG-ului (electromiografie) (Andersen et al 1985) si MRI (scanare prin rezonanta magnetica) (Richardson et al 1998) cat si prin folosirea inregistrarii acumularii de caldura ca si indicator al producerii de energie in diferite parti ale muschiului quadriceps (Krustrup e al 2004), s-a observat ca in timpul exercitilor moderate si intense de extensie a piciorului, 90-100% din efort, activitate musculara si productie de energie este atribuit muschilor coapsei care realizeaza extensia piciorului(Andersen and Saltin 1985; González-Alonso et al 2000). Mai mult, prin umflarea garoului plasat sub genunchi se impiedica contaminarea sangelui care iriga muschii coapsei de catre sangele care vine din gamba.
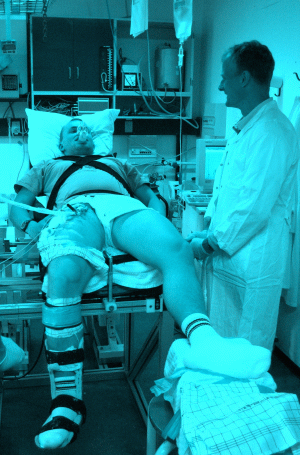
Fig 1. Extensia piciorului din asezat, ca model de exercitiu folosit in investigatiile din cadrul fiziologiei sportive, a fost introdus de Andersen et al (1985). In acest model subiectul este culcat dorsal, cu piciorul care efectueaza extensia atasat la un ergometer electric Krogh or Monark printr-o tija de aluminiu. Modelul a fost imbunatatit in studiul lui Bangsbo et al (1990), in care subiectul a fost plasat in pozitia mentionata, cu scopul de a evita eventualele miscari efectuate de tors si pentru a usura accesul la grupele musculare angajate in efort (i.e. extensorii coapsei).
Alt beneficiu al folosirii acestui exercitiu, este ca zonele active ale quadricepsului sunt expuse in vederea recoltarii biopsiilor musculare (Fig 1). De asemenea rezistenta externa este usor de controlat prin posibiliatea de stabili intensitatea de franare a rotii plasate inapoia scaunului pe care este asezat subiectul. Totusi, din analizele efecuate pe biopsiile musculare este dificil sa estimezi metabolismul energetic in timpul eforturilor fizice desfasurate in timpul pedalarii, alergarii s.a.m.d., deoarece masa si activitatea muschilor angrenati in efort nu este cunoscuta precis si activitatea metabolica a muschilor biopsiati nu poate fi reprezentativa pentru tote grupele musculare recrutate de tipul si intensitatea exercitiului.
O prima abordare indirecta a productiei anaerobe de energie este masurarea deficitului de oxigen determinat de intensitate pe durata exercitiului. Acest termen a fost introdus de Krogh si Lindhard (1920) si reprezinta diferenta dintre consumul de energie estimat si consumul total de oxigen. Se pare insa ca, deficitul de oxigen poate fi folosit in masurarea energeticii anaerobe doar in exercitiile submaximale unde relatia consum de oxigen-lucrul mecanic efectuat este lineara. In cadrul exercitiilor submaximale mari si maximale, asa cum a fost descris si in sectiunea care descrie energetica aeroba, aceasta relatie devine non-lineara (Fig 2, linia intrerupta).
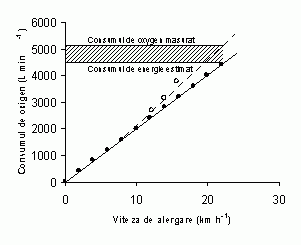
Fig 2. Relatia non-lineara dintre consumul de oxigen si lucrul mecanic efectuat in cadrul eforturilor maximale si supramaximale poate furniza date eronate in folosirea ei pentru calcularea deficitului de oxigen; onsumul de oxigen estimat (linia continua) si consumul de oxigen masurat (linia intrerupta).
In acest caz, consumul de energie determinat prin extrapolarea liniara a relatiei dintre consumul de oxigen-lucrul mecanic efectuat in cadrul eforturilor submaximale va fi subestimat. Aceasta inseamna ca, cu cat intensitatea exercitiului este mai mare cu atat mai pronuntata va fi subestimarea adevaratului deficit de oxigen (Fig 2). Totusi, in studiul lui Bangsbo et al (1990), estimarea productiei anaerobe de energie in extensia piciorului din asezat, unde masa musculara angrenata in efort este redusa si poate fi destul de precis cuantificata, a corespuns deficitului de oxigen.
Masurarea debitul de oxigen (cantitatea de oxigen consumata in perioada de recuperare de dupa terminarea efortului), este o alta cale indirecta de investigare a energeticii anaerobe (Fig 3). Debitul de oxigen poate fi calculat prin extragerea consumul bazal de oxigen (consumul de oxigen in stare de repaus) din consumul total de oxigen masurat de-alungul recuperarii dupa incetarea efortului. In mod normal, rata consumului de oxigen post exercitiu scade la jumatate de-alungul a 30 de secunde (Mc Ardle 2001). Recuperarea post efort este considerata a fi completa cand nivelul consumului de oxigen atinge nivelul acestuia din starea de repaus. Chiar daca laureatul la premiul Nobel Archibald Vivian Hill, a folosit in anul 1922 termenul de debit in ideea platirii datoriei de oxigen facuta la inceputul efortului, deficitul de oxigen, si a presupus o legatura directa intre aceste doua procese fiziologice, se pare ca, absorbtia oxigenului in perioada recuperatorie, post exercitiu, este mai mare decat nevoia de oxigen determinata de refacerea rezervelor de FCr, ATP si glicogenul resintetizat din lactatul acumulat si eliberat in sange, cat si reancarcarea cu oxigen a mioglobinei din muschi si hemoglobinei din sange. Adevarul acestei afirmatii este dovedit de rezultatele multor studii, dintre care cel al lui Bangsbo et al (1990) este destul de elocvent. Ei au descoperit ca, debitul de oxigen in exercitiul de extensie a piciorului din asezat a fost mai mare decat cantitatea de energie produsa anaerob. Se pare deci, ca deficitul de oxigen nu poate fi folosit pentru determinarea energeticii anaerobe nici in modelul de exercitiu care foloseste grupe musculare izolate, nici in eforturi fizice care angreneaza intreg organismul. In figura 3, se poate observa cum eforturile submaximale mari si maximale cauzeaza un debit de oxigen crescut si o mai lunga perioda de recuperare. Dupa eforturile maximale si supramaximale, recuperarea post efort poate dura si pana a doua zi (linia puncata).
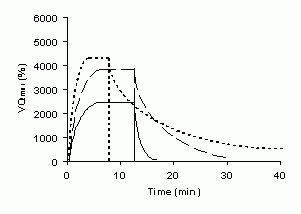
Fig 3. Absorbtia de oxigen in timpul efortului si pe perioada de recuperare dupa efort usor (linia continua), dupa efort moderat (linia intrerupta), dupa efort maximal (linia punctata). In aceasta figura se poate observa ca in efortul maximal unde deficitul de oxigen este cel mai mare, consumul de oxigen in recuperarea post efort nu atinge nivelul celui premergator efortului. (unitatile de masura prezentate in figura sunt orientative).
Exprimarea metabolismului energetic pe baza estimarii energeticii aerobe si anaerobe
Daca producerea de energie pe baza oxigenului consumat a fost discutata in prima parte a acestui articol, productia de energie asociata cu sursele de energie utilizate anaerob (i.e. ATP, FCr si acumularea si eliberarea de lactat care de fapt reprezinta degradarea aneroba a glucozei in cadrul glicolizei), este obtinuta prin converisa degradarii lor in joules folosind valoarea energetica de 55kJ mol-1 de ATP si FCr consumat (Tran et al 1999). Echivalentul pentru producerea de lactat este de 67kJ mol-1 per ATP produs (Curtin et al 1978) in timp ce, pentru resinteza unui mol de ATP in cadrul fosforilarii oxidative sunt necesari 95kJ, la o rata a producerii de fosfati pe consum de oxigen (P/O) de 2.5. Deoarece in conditii standard (0ºC si cantitatea reactantilor de 1 mol) hidrolizarea unui mol de ATP elibereaza o cantitate de energie egala cu 30.5 kJ, poate fi remarcat faptul ca, surplusul de energie eliberat in producerea ATP-ului, creste de-alungul lantului de reactii chimice ce formeaza metabolismul energetic al organismului. Cea mai mare cantitate de energie care nu intra in formarea ATP-ului, are loc in cadrul lantului respirator din mitocondrii (95 vs 30.5kJ per mol ATP).
Productia de energie nu se realizeaza pe seama activarii unei singure cai metabolice (i.e. sistemul fosfagen, glicoliza si fosforilarea oxidativa), ci printr-o interrelationare a acestor sisteme, care este dictata de intensitatea si durata efortului.
In concluzie, poate fi spus ca estimarea directa a consumului de oxigen fie prin masurarea lui la nivelul plamanilor fie prin multiplicarea extractiei acestuia cu viteza de curgere a sangelui, este un bun indicator al energeticii aerobe din timpul efortului. Productia anaeroba de energie este mai dificil de masurat la nivelul intregului organism, dar poate fi destul de precis estimata in eforturi realizate de grupe musculare izolate. Deficitul de oxigen poate fi folosit in predictia energeticii anaerobe in cadrul eforturilor submaximale si probabil in exercitiile efectuate de grupe musculare izolate cum este extensia piciorului din asezat, in timp ce, in eforturile maximale acesta poate produce erori. Masurarea debitului de oxigen nu poate fi folosita in determinarea contributiei energeticii anaerobe la productia totala de energie in timpul efortului fizic.
27 februarie
Bibliografie
Andersen P. and Saltin B. (1985). Maximal perfusion of skeletal muscle in man. J Physiol (Lond) 366:233-249.
Andersen P., Adams R.P., Sjogaard G., Thorbe A and Saltin B. (1985). Dynamic knee extension as a model for the study of an isolated exercising muscle in man. J Appl Physiol 59:1647-1653
Bangsbo J., Gollnick P.D., Graham T.E., Juel C., Kiens B., Mizuno M. and Saltin B. (1990). Anaerobic energy production and O2 deficit - debt relationship during exhaustive exercise in human. J Physiol. 422:539-599.
Bangsbo J., Petersen A. and Michalsik L. (1993). Accumulated O2 deficit during intense exercise and muscle characteristics of elite athletes. Int J Sport Med 14:207-213.
Bangsbo J., Aagaard T., Olsen M., Kiens B., Turcotte L.P. and Richter E.A. (1995). Lactate and H+ uptake in inactive muscles during intense exercise in man. J Physiol (Lond) 422:539-559.
Bangsbo J., Krustrup P., González-Alonso J. and Saltin B. (2001). ATP production and efficiency of human skeletal muscle during intense exercise: effect of previous exercise. Am J Physiol 280(6):E956-E964.
Barclay C.J. (1996). Mechanical efficiency and fatigue of fast and slow muscles of the mouse. J Physiol 497:781-794.
Crow M.T. and Kushmeric M.J. (1982). Chemical energetics of slow and fast-twitch muscles of the mouse. J.Gen Physiol 79:147-166.
Curtin N.A. and Woledge R.C. (1978). Energy changes and muscular contraction. Physiol rev 58:690-761.
Gollnick P.D., Piehl K. and Saltin B. (1974). Selective glycogen depletion pattern in human skeletal muscle fibres after exercise of varying intensity and at varying pedaling rate. J Physiol (Lond) 241:45-57
González-Alonso J., Quistorff B., Krustrup P., Bangsbo J. and Saltin B. (2000). Heat production in human skeletal muscle at the onset of intense dynamic exercise. J Physiol 524 (2):603-615
Hellsten Y., Richter E.A., Kiens B. and Bangsbo J. (1999). AMP deamination and purine exchange in human skeletal muscle during and after intense exercise. J Physiol (Lond)520:909-919.
Katz A., Broberg S., Sahlin K. and Wahren J. (1986). Muscle ammonia and amino acid metabolism. Clin Physiol 6:365-379.
Krogh A. and Lindhard J. (1920). The changes in respiration at the transition from work to rest. J Physiol 53:431-437.
Krustrup P., Sönerlund K., Mohr M., González-Alonso J. and Bangsbo J. (2004). Recruitment of fiber types and quadriceps muscle portions during repeated, intense knee-extension exercise in humans. Pflügers Arch 449(1):56-65.
McArdle W.D., Katch. F.I. and Katch V.L. (2001). Exercise Physiology: energy, nutrition and human performance/ Fifth edition. 2001
Pedersen P.K., Sørensen J.B., Jensen K., Johansen L. and Levin K. (2002). Muscle fiber type distribution and nonlinear VO2-power output relationship in cycling. Med Sci Sports Exerc 34(4):655-661
Pringle J.S., Doust J.H., Carter H., Tolfrey K. and Jones A.M. (2003). Effect of pedal rate on primary and slow-component oxygen uptake responses during heavy-cycle exercise. J Appl Physiol 94(4):1501-1507.
Richardson R.S., Frank L.R. and Haseler L.J. (1998). Dynamic knee-extensor exercise: functional MRI of muscular activity. Int J Sports Med 19(3):182-187.
Spriet L.L., Sőnderlund K., Bergstrøm M. and Hultman E. (1987). Anaerobic energy release in skeletal muscle during electrical stimulation in men. J Appl Physiol 62:611-615.
Tran T.K., Sailasuta N., Kreutzer U., Hurd R., Chung Y., Mole P., Kuno S. and Jue T. (1999). Comparative analysis of NMR and NIRS measurements of intracellular PO2 in human skeletal muscle. Am J Physiol Regulatory Integrative Comp Physiol 276:R1682-R1690.
Vøllestad N.K. and Blom P.C. (1985). Effect of varying exercise intensity on glycogen depletion in human muscle fibers. Acta Physiol Scand 125 (3):395-405
Willis W.T. and Jackman M.R. (1994). Mitochondrial function during heavy exercise. Med Sci Sports Exerc 26:1347-1354
Woledge R.C. (1998). Techniques for muscles energetics. Current Methods in Muscle Physiology. ed. Sugi H, Oxford University Press
Zolads J.A., Rademark A.C. and Sargeant A.J. (1995). Non-linear relationship between O2 uptake and power output at high intensities of exercise in humans. J Physiol 488(1):211-217.
Zunts N. (1901). Ueber die Bedeutung der verschiedenen Nährstoffe als Erzeuger der Muskelkraft. Arch Gesamte Physiol, Bonn, Germany: LXXXIII: Pflűgers Arch Physiol; 83:557
|
