Studii si cercetari privind formele de manifestare ale vitezei in proba de sprint
Specialistii domeniului gasesc, pentru viteza, mai multe modalitati de manifestare, determinate in mare masura de modalitatea in care acestea intra in compozitia miscarilor probelor de sprint si de factorii ce o influenteaza.
In ceea ce priveste viteza de alergare din proba de sprint, V. Filin, 1978, deosebeste:
Rapiditatea reactiilor motrice;
Capacitatea de a produce frecvente 343g63d maxime ale miscarii;
Capacitatea maxima de accelerare;
Capacitatea de a atinge si de a mentine maximum de viteza intr-un anumit spatiu.
Aceasta interpretare a vitezei de alergare justifica partial clasarea vitezei printre aptitudinile conditionale.
C. Capizzi 1978, citat de R. Manno, 1996, defineste viteza numai prin componentele ei elementare. Manno, 1996, asimileaza viteza cu urmatoarele forme de rapiditate: rapiditate de reactie, rapiditate de repetitie, rapiditate de deplasare, rapiditate de rezistenta.
Vom trata in continuare formele de manifestare ale vitezei care intra in componenta probei clasice de sprint, 100 mp, sub denumirea lor obisnuita, asa cum apar in publicatiile romanesti.
Viteza de reactie
Viteza de reactie se substituie de fapt cu timpul de reactie.Timpul de reactie motrica, in conceptia lui R. Manno, 1996, are o componenta perceptiva, de organizare a miscarii, dar fara factori limitativi de tip energetic, exceptand situatiile deosebite ca: oboseala, repetari numeroase, rezistente mari de invins. J. Weineck, 1993, considera ca acest timp se subordoneaza legilor fiziologice senzoriale, care nu-i permit sa scada sub valoarea limita de 1/10s.
M.
Pradet, 1997, denumeste viteza de reactie ca fiind capacitatea
sportivului de a percepe, de a analiza si de a prelucra in timpul cel mai
scurt posibil, semnalul care declanseaza actiunea, astfel incat
aceasta sa debuteze cu o pierdere minima de timp.
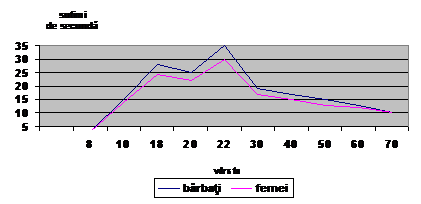
B. A. Podlovaev, 1979, sustine ca cel mai inalt tempou de dezvoltare a reactiei vizual motrice se inregistreaza la varsta cuprinsa intre 8-12 ani. M. Grosser si S. Rostislav, 1976, citati de T. Ardelean, 1991, sustin ca timpul de reactie scade intre varsta de 8-25 ani, sub influenta actului motric specific, se stabilizeaza pana la 60 de ani, dupa care creste.
|
Varsta | |||||||||||||
|
Timp de reactie |
| ||||||||||||
|
Viteza de miscare pura | |||||||||||||
|
Viteza de miscare contra rezistenta |
| ||||||||||||
|
Frecv. contactelor | |||||||||||||
|
Viteza | |||||||||||||
|
Frecventa miscarii |
Tabel nr.3. Perioadele sensibile pentru dezvoltarea vitezei.
Dupa Cometi,1989, citat de Gaçon,1995.
R. Manno, 1996, considera ca timpul de reactie se imbunatateste foarte mult, in prima etapa intre 6(8)-18 ani, apoi creste pana la 20 ani. Intre 20-22 ani, acelasi timp cunoaste o evolutie buna, dupa care creste din nou pana dupa varsta de 70 ani.
Varsta optima pentru imbunatatirea timpului de reactie este, pentru majoritatea specialistilor, 6½ -10 ani. In conceptia lui Gaçon, 1995, timpul de reactie se imbunatateste cel mai bine in perioada varstei de 6-10 ani.
In conceptia lui J. Weineck, 1983, viteza de reactie are mare importanta in probele de sprint si este putin perfectibila, avand un ritm de dezvoltare propriu, in care etapa de varf se situeaza in jurul varstei de 20 ani. Tot J.Weineck, 1983, considera ca o prima etapa de imbunatatire a timpului de reactie exista in jurul varstei de 10 ani. Apoi timpul de reactie scade de la 500-600 la 200-400 miimi de secunda, ca la 14 ani sa atinga aceleasi valori ca ale adultului.
In alta ordine de idei privind timpul de reactie la startul de jos, G. Dintiman, B.Ward si T. Tellez, 1997, sustin ideea ca, daca atentia este concentrata pe pocnetul pistolului, apare o reactie mai inceata, deoarece trebuie sa-ti muti atentia de pe sunet pe miscare.
Atentia se concentreaza mai mult pe prima miscare musculara, ca de exemplu actiunea rapida a bratelor sau o ridicare rapida a genunchiului, decat pe sunetul pistolului. Oricum se aude pocnetul unui pistol de calibru de 22, de la 2m departare, fara sa fie nevoie sa asculti acest sunet.
E. Plocon si D.Alexandrescu,1980, in urma unui studiu realizat pe finalistii J.O. de la Moscova, au evaluat timpul de reactie a sprinterilor in proba de 100mp, stabilind treptele valorice ale timpului de reactie la startul cursei de sprint, in speta proba de 100mp.
Vom prezenta prin comparatie cu studiul lui E. Plocon si D. Alexandrescu, 1980, rezultatele finalistilor de la proba de 100mp, de la J.O. de la Sydney din 2000 in tabelul de mai jos.
|
Calificativ |
Timpul de reactie (s) | ||
|
exceptional | |||
|
foarte bun | |||
|
bun | |||
|
mediocru | |||
|
slab | |||
|
foarte slab |
peste 0,230 |
peste 0,260 |
|
Timpul de reactie la JO de la Sydney este incomparabil superior pentru primele trei calificative. Restul de calificative au valori nesemnificativ apropiate. In ceea ce priveste raportul intre timpul de reactie si rezultatul final in proba de sprint 100mp, aceiasi autori au demonstrat ca nu exista nici o corelatie.
Tot din studiul lui E. Plocon si D. Alexandrescu, 1981, aflam ca intre timpul de reactie si marimea distantei de alergare din probele de sprint exista un raport direct proportional: cu cat creste distanta cu atat este mai mare valoarea timpului de reactie.
Viteza de accelerare
Nivelul relativ al vitezei de reactie, vitezei de executie si vitezei de repetitie, in speta cei trei parametri asociati nivelului de forta musculara pentru a invinge inertia, alcatuiesc viteza de accelerare a fiecarui sprinter.
In conceptia lui T. Ardelean, 1991, dezvoltarea acestei forme de manifestare a vitezei, se bazeaza pe dezvoltarea simtului de accelerare si duce la cresterea capacitatii de accelerare.
Aceasta nu se combina obligatoriu cu acea capacitate care permite mentinerea unei viteze ridicate de deplasare. De aici deriva si necesitatea de a lucra separat aceste doua componente.
In atletism, posibilitatea de a putea mari in mod rapid, cadenta alergarii si a deplasarii cu o viteza ridicata este relativ independenta. (M.V. Zatiorski, 1972). In aceeasi ordine de idei, G. Harbin, 1989, a constatat ca antrenamentul poate imbunatati sincronizarea miscarilor, ceea ce ar duce la imbunatatirea timpului de miscare pe seama imbunatatirii coordonarii, dezvoltarii fortei rapide, imbunatatirii elasticitatii si mobilitatii, perfectionarii tehnice.
Profesorul L. Seagrave, in 1996, a relevat faptul ca timpul de accelerare depinde de urmatoarele componente:
* timpul de reactie;
*in startblocuri, prin antrenament, forta de propulsie a picioarelor depinde de nivelul de forta si puterea exploziva.
*viteza de miscare a bratelor/picioarelor.
Fetele ating platoul maxim de dezvoltare a frecventei la varsta de 13-16 ani, in timp ce baietii ating la varsta de 18 ani.
Cand atletul doreste sa-si accelereze greutatea corpului in deplasare, cu viteza si frecventa mare a pasilor in probele de sprint, acest fapt se datoreste capacitatii lui de a exprima varfuri inalte de forta in timp scurt si tot mai scurt.
De aceea, una din caracteristicile vitezei este tocmai aceea ca ea este constituita din parametri relativ independenti unii fata de ceilalti, implicand o dezvoltare generala care necesita utilizarea unor proceduri foarte variate.
Astfel ca, daca s-a reusit sa se accelereze pana la atingerea vitezei maxime in cursa de sprint, viteza maxima nu poate tot atat de bine sa fie mentinuta pe o durata considerabila. Farfel citat de R. Manno, 1996, exemplifica evolutia frecventei maxime de miscare in functie de varsta si sex. Astfel, intre 14 -16 ani, fetele obtin acceasi frecventa maxima, de 5,2 miscari pe secunda, in timp ce baietii la varsta de 14 ani, au frecventa maxima de miscare, de 5,4 miscari pe secunda, care creste pana la 16-17 ani la 5,6 miscari pe secunda. G. Cometti, 1989, sustine ideea ca, intre 7 13 ani, viteza de accelerare (frecventa miscarii) va avea prioritate. Pe pista de alergare, frecventa miscarii se exprima prin numarul de fuleuri in unitatea de timp, iar dezvoltarea lor pare a fi optima intre 9 11 ani, cand atinge valori de 4,44 pasi / secunda la baieti si 4 pasi / secunda la fete, (fara a se specifica in vreun studiu si lungimea fuleului corespunzator acestei frecvente, n.a.) dupa care scade usor pana la 15 16 ani.
Fiziologii, biologii si teoreticienii antrenamentului sportiv ca: R. Malina, C. Bouchard, 1991, Frey, 1993, citat de R. Manno, 1996, sustin ca varsta de 10 -13 ani este propice pentru dezvoltarea frecventei, apoi scade brusc pana la 17 ani, dupa care din nou se amelioreaza. L. Seagrave, 1996, crede ca, pentru a nu fi afectata frecventa optima intr-o cursa de sprint, lungimea fuleului poate sa fie mai mare cu maxim 15cm.
O crestere a fuleului de 15 cm duce la o crestere a vitezei de 0,6m.s-1. Urmarea acestui fapt este ca in fiecare secunda se poate parcurge o distanta suplimentara de 60 cm. Pe distanta parcursa in 10 s-ar economisi timpul parcurs pentru aproape 6m. Deci, cursa de 100 m se parcurge cu 0,5-0,9s mai repede.
Acelasi autor conchide si alte aspecte, ca:
- frecventa fuleului este data pe de o parte de suma timpilor de contact cu solul si timpilor de stat in aer. Pentru sprinterii campioni raportul este de 2/1 la startul cursei si devine 1/1,3-1/1,5 la maximum de viteza, in partea a doua a cursei;
- pe de alta parte, frecventa fuleului este determinata de viteza, unghi, inaltimea saltului si rezistenta aerului.
- femeile sunt cu aproximativ 1s mai lente decat barbatii in proba de 100 mp, datorita frecventei mai reduse. Principala cauza o constituie diferentele de forte de reactie la contact cu solul si de putere.
Lungimea efectiva a pasului poate fi imbunatatita prin trei metode, recomandate de L. Seagrave, 1996:
a. marirea capacitatii de efort a atletului la nivelul pistei, prin imbunatatirea fortei si puterii atletului;
b. marirea efortului aplicat prin perfectionarea tehnicii;
c. reducerea frecarii de tragere sau de franare, rezultate la aterizarea fiecarui pas de alergare; tragerea este determinata de modul cum atletul aseaza piciorul la contactul cu solul. Frecarea din interiorul pantofului de atletism (cuie) reprezinta un alt tip de influenta.
Frecventa miscarii, prin care se exprima de fapt viteza de repetitie, isi gaseste reflectare in toate miscarile ciclice si se bazeaza permanent sau cu regularitate pe o aceeasi schema gestuala. In probele de sprint o regasim in frecventa pasilor de alergare, ca fiind o componenta a capacitatii de accelerare si a vitezei de deplasare.
Despre capacitatea de accelerare, G. Harbin, 1989, sustine ca ea depinde de capacitatea sistemului nervos central de a emite rapid o succesiune de stimuli nervosi, pentru ca forta sa se manifeste rapid.
Ea incepe cand timpului de reactie ii urmeaza timpul de miscare, sustine acelasi autor. Timpul de miscare poate fi imbunatatit numai prin limitarea valorii repetarilor pentru o sarcina specifica, altfel spus, cu cat repeti mai mult miscarea pentru o anumita sarcina si cu o viteza aproape de maxim, cu atat timpul de miscare va fi mai scurt (dar in acelasi timp avut grija sa nu se instaleze bariera de viteza, n.a.)
Spre deosebire de tehnica de obtinere a vitezei maxime, in tehnica de accelerare, in conceptia lui F. Urtebise, 1994, talpile atletului vor fi pozitionate in spatele proiectiei centrului de masa pentru primii pasi de alergare de dupa start. Cu fiecare pas suplimentar prin care sprinterul castiga experienta, distanta dintre proiectia centrului de masa si baza de sprijin se va micsora inainte ca talpile sa aterizeze direct sub trunchi. In aceasta faza a cursei, directia fortei aplicata este mai mult orizontala.
Acelasi autor releva faptul ca o tehnica corecta de accelerare necesita ridicarea genunchilor cu rapiditate la nivelul maxim. Unghiul dintre gamba pamant si trunchi pamant vor fi egale. Glezna este indoita inaintea contactului puternic cu pamantul.
F. Urtebise, 1994, sustine ca suprafata sintetica pe care se alearga astazi impune o noua tehnica de alergare si mai ales invatarea de catre copil a impulsiei pe suprafete artificiale. Despre acest fapt, autorul evidentiaza urmatoarele aspecte:
*eficacitatea impulsiei unui atlet depinde de modul cum sistemele lui fiziologice contribuie cu energia elastica si chimica necesara in alergarea de viteza.
*pe suprafata sintetica, atletul trebuie sa tasneasca ca o rigla metalica si nu ca un baton de plastilina; in noua conceptie de rigla metalica, articulatia gleznei, genunchiului, soldului, umarului sa se gaseasca in aceeasi linie in axul de contact cu solul.
*pentru ca impingerea in sol sa fie eficace, atletul este obligat sa deschida mai mult gamba spre inainte pentru a amplifica agatarea solului printr-o miscare dinainte inapoi pe directia de deplasare; deschiderea gambei inainte, ridicarea genunchiului la orizontala, flexia labei pe gamba determina realizarea unui timp de cuplare scurt, excentric / concentric al contractiei.
F. Urtebise, 1994, considera ca aceasta miscare pentru inaintare care este rezultanta impingerii, nu mai poate fi numita impingere ci intindere.
In acest caz exista 3 tipuri de intinderi:
1*intindere aeriana - cand membrul inferior se afla in perioada de oscilatie;
2*intindere de tensiune - cand incepe faza de contact;
3*intindere de transmitere - propulsia propriu- zisa, astfel faza de impulsie este mai mult o tractiune-intindere decat impingere-intindere (n.a), adica intindere aproape completa orientata inainte cu o mai puternica componenta a fortei orizontale ca actiune si implicit si reactiune. F. Urtebise, 1994.
In alta ordine de idei, puterea (mecanica) este un parametru biomecanic important atunci cand analizam performantele alergarii umane, deoarece aceasta semnifica eliberarea de energie care initiaza sau controleaza miscarea fiecarui segment. In dinamici diverse, momentul fiecarei articulatii este determinat de combinatia dintre cinematicile segmentare si articulare, masurile antropometrice si fortele externe.
Dupa cum am mentionat in subcapitolele anterioare, rezultatele obtinute in probele de sprint sunt determinate in cea mai mare parte de viteza de accelerare. Rezultatele valoroase ale sprinterilor de nivel mondial si olimpic sunt si urmarea aplicarii in practica de antrenament a cercetarilor minutioase a specialistilor din domeniul fiziologic, biomecanic si biofizic la nivelul musculaturii. Pasul alergator de accelerare a fost si este foarte minutios analizat, astfel incat fiecare cercetator a contribuit la redarea fidela a alergarii in timpul fazei accelerarii, pentru a determina puterea mecanica necesara atletului in alergare. Printre cercetatorii acestor probleme, enumeram pe cei care au studiat mai recent: P. Astrand si K. Rodahl, 1986, M.J.L. Alexander, 1989, V. Vardaxis si T. B. Hoshizaki, 1989, D.A. Winter, 1990, R. Jacobs si Van Schenau, 1992, R.V. Gülch, 1994, C.H. Delecluse, 1997, U.M. Kujala si M. Järvinen, 1997, M. J. L. M. Dowson et al, 1998, M. D. Johnson si G. Buckley, 2001.
D.A. Winter, 1990, sustine ca, puterea mecanica poate sa fie calculata ca fiind produsul dintre momentul articular si viteza unghiulara a articulatiei si poate oferi informatii despre a activitatea tuturor muschilor la nivelul fiecarei articulatii.
Aceasta cale a fost utilizata pentru a determina actiunea (fie ea excentrica sau concentrica) a grupurilor de muschi (flexori sau extensori) care intrepatrund articulatia.
V. Vardaxis si T. B. Hoshizaki, 1989, citati de M. D. Johnson si G. Buckley, 2001, au cercetat care este puterea mecanica a muschilor extensori si flexori ai membrului inferior, nu numai in faza de sprijin ci si in perioada de pendulare, (pas anterior, pas posterior) in timpul alergarii de accelerare.
Studiul a fost realizat cu 6 atleti de nivel mediu care au sprintat pe o platforma cu senzori.
Mai exact, acestia au cercetat dinamica momentului de forta a articulatiei soldului, genunchiului si gleznei in Nm, (tab. nr. 4) si a puterii musculare in timpul acestor faze inW
In tabelul urmator (dupa aceiasi autori) sunt prezentate valorile vitezei unghiulare (rad.s1), a momentelor de varf ale acelorasi articulatii (Nm) si puterii musculare(W) pentru sold(H), genunchi (K) si glezna (A) a fiecarui sprinter, impreuna cu o medie omogena a grupului de sprinteri.
|
articulatie |
pasul posterior |
sprijinul |
pasul anterior |
|
Hip ( sold) |
*HM1:-275±42 (extensor) |
HM2: 280±156 (flexor) HM3:-377±34 (extensor) |
HM4: 94±181 (flexor) HM5:281±61 (flexor) |
|
Knee (genunchi) |
**KM1:-135±18 (flexor) |
KM22 269±106 (extensor) KM3: 231±90 (extensor) |
KM4: 41±8 (extensor) |
|
ankle (glezna) |
neglijabil |
*** AM1: -328±89 (extensor) |
neglijabil |
* momentul de varf al soldului * momentul de varf al genunchiului,*** momentul de varf al gleznei (N.m)
Tabel nr.4. Momentele de varf ale articulatiilor. V. Vardaxis si T.B. Hoshizaki, 1989, citati de D.M. Johnson si G.J. Buckley, 2000.
In paginile urmatoare vom prezenta cum lucreaza flexorii si extensorii celor 3 articulatii in 3 faze: dupa faza de mijloc a balansului pana la pasire, la pasire, in pasul posterior in timpul alergarii accelerate din faza de tranzitie a acceleratiei. Vom prezenta pe rand aspectele esentiale ale acestor faze:
A) Dupa faza de mijloc a balansului pana la pasire
Soldul initial flexat se extinde. Puterea musculara se schimba de la (-) la (+) : de la HP1 la HP2 atingand valoarea maxima de 2179±328W cand muschii extensori lucreaza de la excentric la concentric.
Genunchiul inainte de pasire este flexat (de la balans pana la pasire) si ii corespunde o putere negativa de - 2097±295 W, prin actiunea extensoare a flexorilor
Glezna in aceeasi faza, puterea mecanica a ei este neglijabila.
B) La pasire
Soldul continua sa se extinda, rezultand o putere flexoare negativa (HP3). In acest timp, forta de reactie a solului isi schimba sensul, ceea ce conduce la extensia puternica a soldului si puterea atinge valoarea maxima de 3242±1086W, cand extensorii lucreaza concentric.
Genunchiul la pasire, viteza unghiulara este minima si corespunde unei puteri neglijabile. Dupa faza mijlocie a pasirii, genunchiul se extinde pana la ridicarea varfului de pe sol, cand se manifesta o putere pozitiva, concentrica a muschilor extensori de 1544±512W.
Glezna - la inceputul fazei de sprijin se afla in flexie plantara prin actiunea extensoare a muschilor flexori. Puterea este negativa si are valoarea de -2205±812W. Dupa mijlocul fazei de sprijin, actiunea flexorilor plantari, se schimba de la excentric(-) la concentric (+), cand puterea este de 3066±846W, propulsand corpul inainte in faza de zbor.
Aceste rezultate sugereaza ca glezna trece printr-un ciclu de intindere-scurtare, unde se stocheaza energie elastica. Puterea maxima a flexiei plantare este cea mai mare putere generata dintre toate articulatiile pentru un pas complet de alergare si singura putere apreciabila generata in ultima faza a sprijinului. Aceasta este de fapt o prima concluzie a studiului. Actiunea gleznei, in generarea vitezei de accelerare, se materializeaza in antrenamentul actual, prin exercitii ca: sarituri ca mingea executate din glezna, schipurile inalte.
C. Pasul posterior
Flexorii soldului lucreaza concentric, generand o putere mare pozitiva de +1699±215W.
Extensorii genunchiului lucreaza excentric generand o putere de -859±256. Puterea generata de glezna este neglijabila.
Concluzia finala a studiului este ca extensia concentrica a soldului din pasul anterior si prima faza de sprijin, extensia concentrica a genunchiului dupa faza de mijloc a sprijinului (KP2) si flexia concentrica plantara din ultima faza a sprijinului (AP2), servesc pentru a genera viteza de sprint in faza de mijloc a accelerarii (de tranzitie).
R. Jacobs si Van Schenau, 1992, au relevat aceste aspecte accentuand urmatoarele:
* Actiunile muschilor biarticulari, contribuie la transferul de putere de la articulatiile proximale la cele distale.
*Actiunea muschilor mari, monoarticulari ai soldului este transferata la glezna si sol, determinand o productie mare de putere care sa ajunga la articulatiile distale, in ciuda faptului ca ele au muschi relativ mici.
Privind efectele exercitiilor excentrice, intr-o recapitulare referitoare la tratamentul si prevenirea accidentelor coapselor, U. M. Kujala si M. Järvinen, 1997, au subliniat faptul ca muschii coapsei sunt accidentati mai ales in timpul exercitiului excentric, cand muschiul capata o tensiune mare in timpul alungirii.
Actiunea piciorului descrisa mai sus poate astfel sa ofere un raspuns referitor la ,,de ce accidentele de coapsa sunt comune in sprint.
Totusi, datorita puterii extensoare pozitive, care a fost generata simultan la sold, e posibil ca muschii posteriori ai coapsei sa actioneze intr-o maniera quasi-izometrica.
Pentru o intelegere mai buna a rolului dublu, pe care il au muschii posteriori ai coapsei, in sprint, sunt necesare mai multe cercetari in viitor, pentru analiza cinematica a fiecarui pas din faza de accelerare pana la mentinerea vitezei maxime.
C.H. Delecluse, 1997, in studiul sau demonstreaza ca dupa 10m de alergare (intr-un sprint la o viteza maxima) rolul muschilor extensori ai genunchiului, de a mentine sus centrul de greutate al corpului, este esential. In acest studiu, el sustine ca, de la pasirea pe sol a piciorului pana la faza mijlocie a sprijinului, este indicat un rol similar al genunchiului prin generarea unei puteri mecanice minime. Numai dupa ce centrul de greutate s-a mutat anterior, pe sprijinul membrului inferior, se genereaza o putere extensoare pozitiva de 1544±512W. Generand astfel puterea, inseamna ca, corpul este impins inainte, nu in sus. Aceasta putere a fost mai mare decat cea raportata de R. Jacobs si Van Schenau, 1992, marimea puterii flexorului plantar din studiul lui V. Vardaxis si T.B.Hoshizaki, 1989, (3066±846W9) a fost comparabila cu cea raportata in 1992 de R. Jacobs si Van Schenau, (≈2180W) si constata ca rolul flexorilor plantari in generarea vitezei orizontale, e important in ambele faze, atat in faza de start cat si in accelerarea de tranzitie. Puterile musculare ale soldului si genunchiului in timpul balansului au fost similare in marime cu cele raportate de V.Vardaxis si T.B.Hoshizaki, 1989, in alergarea cu viteza maxima.
Nu se stie inca cu exactitate daca miscarile articulare asociate cu faza de mijloc a accelerarii, sunt similare cu acelea asociate cu ambele faze: startul si viteza maxima; comparatiile acestea sunt limitate deoarece nu s-au gasit date referitoare la puterea soldului, genunchiului si gleznei, din faza de sprijin din alergarea cu viteza maxima. Aceste descoperiri se aduna la masa de cunostinte privind biomecanica sprintului, asigurand o certitudine in actiunea membrului inferior, in generarea vitezei din timpul tranzitiei dintre start, accelerare si obtinerea vitezei maxime.
Cei doi cercetatori conchid ca pentru o intelegere mai buna a contributiilor articulare ale membrului inferior la obtinerea vitezei maxime, studiile viitoare ar trebui sa incerce sa determine puterile musculare de-a lungul fiecarui pas, consecutiv, de la plecarea din startbloc pana la obtinerea vitezei maxime. Cu aceasta ocazie se consemneaza ca puterea musculara este un factor major in succesul atletului in probele de sprint. Pentru a intelege cum viteza acumulata in faza de accelerare este obtinuta si mentinuta in faza de viteza maxima, e necesar sa determinam care sunt parametri biomecanici implicati si in urmatoarea faza.
M.J.L. Alexander, 1989, citat de M. Dowson si altii, 1998, a studiat sprinteri de elita si a gasit corelatii semnificative intre timpii la sprint pe 100 m si varful de forta a contractiei concentrice a genunchiului in extensie. El a sugerat ca, teoretic, performanta la sprint este rezultatul direct al produsului fortei medii si timpului de contact aplicat de atlet impotriva solului, in timpul fazei de propulsare a pasului anterior.
Forta conducatoare generata in timpul fazei de aterizare este legata de puterea in flexie si in extensie a soldului, de extensia genunchiului si flexiunea plantara.
De aceea, s-ar parea ca atletii, care au o putere mai mare in aceste grupe de muschi, vor scoate la alergare de viteza timpi mai buni, daca timpul de contact ramane acelasi.
B. Nielson si altii, 1980, au studiat efectele a trei moduri de antrenament pe grupe de fete, cu varste cuprinse intre 7-19 ani. Fetele au fost impartite in grupe dupa inaltime.
Grupele antrenate au fost pregatite astfel: forta izometrica-24 extensii maximale ale genunchilor pe antrenament, antrenament de alergare-100 de pasi in 10 starturi pe sedinta de antrenament, saritura verticala-80 sarituri pe sedinta. Numai grupa antrenata izometric a demonstrat o crestere semnificativa a fortei izometrice de extensie voluntara a genunchiului comparativ cu grupele de control si numai grupa antrenata cu sarituri si-a imbunatatit performantele la saritura pe verticala.
M. Dowson, 1998, a realizat un studiu cu participarea unui grup de 8 atleti care au alergat pe distanta 0-15 m, 0-30m, 0-35 m fiind inregistrati electronic cu celule fotoelectrice. S-a masurat puterea izokinetice a articulatiilor glezna, genunchi, sold, prin comparatie cu alti 8 sportivi in activitate dar care nu erau atleti.
Datele antropometrice ale celor doua grupe de sportivi sunt:
varsta talia(cm) greutatea(kg) lung. tibiei(cm)
atleti 22,5± 3,7 180,8±5,4 77,3±8,3 41,6±1,6
alti sportivi 28,5±8,4 180,1±7,5 75,5±6,5 42,7±2,7
Cei 16 sportivi au alergat distanta de 15m trecand fiecare prin fata a 4 celule fotoelectrice, plecarea fiind cu start de jos. Apoi au alergat distanta de 35m, trecand prin fata primei celule fotoelectrice asezata la distanta de 30m iar a doua celula la 35m.
Un Cybex de forta a fost folosit ca instrument de testare valid si de incredere pentru fiecare grupa musculara: flexorii si extensorii soldului, ai genunchiului, flexorii si dorsoflexorii plantei de la glezna piciorului dominant. Toate actiunile au fost testate la viteze mici, medii si mari. Fiecare subiect a fost masurat pe un scaun adaptabil, trunchiul fiind imobilizat prin curele. La fiecare test de velocitate, subiectul a efectuat trei incercari de incalzire submaximale urmate de alte trei incercari maximale. Din acest studiu, a rezultat faptul ca timpul pe distanta 30-35m nu este puternic influentat de flexorul concentric al gleznei si extensorul concentric al genunchiului. In tabelul de mai jos, vom prezenta rezultatele studiului.
|
0-5 m | ||
|
5-10 m | ||
|
10-15 m | ||
|
0-15m | ||
|
30-35m |
*sprinteri; ** alti sportivi, o- p< 0,05
Tab. nr.5. Timpul sprintului(s) pe fiecare interval. Dupa M. Dowson si altii, 1998.
Cea mai mare forta a fost obtinuta de atleti, in actiunea excentrica a genunchiului realizata la viteza cea mai mare (p< 0,05).
Cea mai mare corelatie a fost gasita intre limita flexorului si extensorului genunchiului in actiune concentrica si timpul obtinut pe distanta de alergare de 30-35m. Limita extensorului concentric al genunchiului (4,19 rad/s) a fost gasita ca fiind cea mai buna in corelatia cu lungimea membrului, masa corpului si timpul alergarii r2 =0,771 si S=0,09.).
Concluziile acestui studiu sunt urmatoarele :
*relatia dintre puterea izocinetica si acceleratie a fost gasita ca fiind putin mai mare(r=0,619 la p< 0,01) decat cea dintre aceeasi putere si performanta alergarii de viteza;
*din tab. nr. 5 reiese faptul ca timpul atletilor pe distanta de 0-15 m cunoaste evolutia cea mai mare, comparativ cu ceilalti sportivi; autorul explica acest fapt prin influenta extensorului genunchiului;
*limita puterii mecanice in muschii extensori ai genunchiului masurata la 4,19 rad/s a dat cea mai buna indicatie pentru actiunile membrului inferior si vitezele testate pentru performanta acceleratiei pe distantele testate. In aceeasi ordine de idei, R.V. Gülch, 1994, citat de M. Dawson si altii, 1997, considera ca viteza creste cand scade forta pentru actiunile concentrice ale muschilor. Pentru actiunile excentrice exista o crestere initiala a tensiunii la viteze mici, fapt ce se datoreaza proprietatilor elastice in interactiunea filamentelor de actina si miozina. In antrenamentul atletic, antrenorii gresesc cand se concentreaza asupra rezultatului interactiunii dintre capacitatea de accelerare si capacitatea de a mentine viteza maxima acumulata si mai putin pe dezvoltarea puterii acestor grupe de muschi.
|
